Metasystem Transitions in Biology
Alexei Sharov
1. Duplication and control
"The metasystem transition creates a higher level of organization, the metalevel in relation to the level of organization of the subsystems being integrated"(Turchin 1976).According to Turchin (1976), metasystem transition requires the following 2 steps:
- Duplication of the original system, and
- Establishment of control over multiple copies.
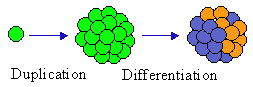
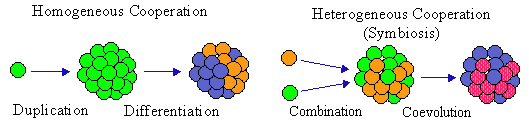
In this figure, the initial element duplicates, then differentiation follows. Differentiation is a typical (but not necessary) result of control of elements by the entire system. However, control always changes system components in order to increase the performance of the entire system.
2. Biological examples
2.1. Hypercycle. The idea of a hypercycle was developed by Eigen and Schuster (1979). They noticed that biological organization results from a cooperation of several autocatalytic systems that augment each other.
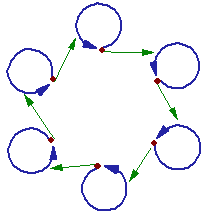
Each autocatalytic component benefits from forming a hypercycle because cooperation increases its reproduction rate. The evolution of hypercycles was never observed experimentally. Thus, the only evidence comes from mathematical simulations. Eigen and Schuster (1979) found that homogeneous systems with uniform components may be evolutionary unstable. They differentiate and a new form (e.g., dissipative structures) emerges at the level of the entire hypercycle. Thus, hypercycles may develop by duplicating of the original gene and subsequent differentiation of genes. This process corresponds well to the Turchin's scheme of metasystem transition.
2.2. Volvox. Volvox (Phytomastigophora) is a colonial algae (diam. up to 1 mm) shown on this picture:
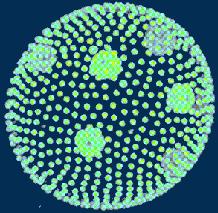
Volvox produces daughter colonies inside which are clearly visible in movies presented at the Microscopy-UK web page. Probably, protection of daughter colonies from predation is one of the functions of the colony.
2.3. Embryogenesis. Embryonic development of most multicellular organisms goes through the stage of blastula which is a hollow ball of cells that resembles a colony of Volvox. For example, below are photographs of early development of sea urchin taken from the web page of Sea Urchin Fertilization Lab

Cells are not differentiated in early embryos. Any cell if separated, can develop into a full organism. This is the stage of multiplication. Differentiation (control) occurs later. At the moment of fertilization, the cell membrane becomes depolarized. This process creates a gradient which later controls the differentiation of cells. The top side of the blastula eventually develops into the ectoderm and the bottom side develops into the endoderm. Further development involves more complicated processes such as gastrulation. More information on embryogenesis can be found at the Frog Embryology web page.
A similar process of multiplication and differentiation occurred in the evolution of segmentation in invertebrates. Worms (Annelida) have almost even segmentation; additional segments appear as worms grow. In most primitive worms, each segment has all organs of the whole organism: paired appendages (parapodia), gut, nervous system, excretory system, reproductive system. Apparently segmentation originated from unfinished vegetative reproduction (parabiosis). Further evolution of segmented worms led to specialization of segments. Earthworms have special segment that produce slime, and only few segments have reproduction functions (see worm anatomy).

Arthropods originated from worms by further specialization of segments. In insects, segments are highly specialized (see Insect Anatomy). For example, 3 thorax segments have legs, 2 of them also have wings. The head developed from several segments that are hard to distinguish. Check the web page on mechanisms of differentiation of segments and Dynamic development.

Adult of the onion fly
Development of insects recapitulates the metasystem transition that occurred at the evolutionary time scale. Many insect larvae resemble worms with a uniform segmentation.

Larvae of the onion fly
2.4. Neural nets. Unicellular organisms can control their functions (movement, feeding, metabolism) mostly via the network of chemical reactions. Because their body is small, diffusion occurs sufficiently fast to transfer signals from one part to another. Other routs of signal transmission (e.g., electrical) are also possible.



 Biosemiotics Homepage
Biosemiotics Homepage