Analysis of Meyen's Typological Concept of Time
Alexei A. Sharov
|
|
Analysis of Meyen's Typological Concept of TimeAlexei A. Sharov
|
|
|
Sergei Meyen was a paleobotanist with a keen interest in philosophy, theoretical biology, and evolutionary theory. His teacher was a famous Russian biologist Alexander Liubishchev. Sergei was a close friend of Julius Shreider, mathematician. Together they generated very interesting ideas in the theory of classification and evolution, and inspired many young (at that time) people including me. Below find my paper on Meyen's "typological concept of time" which was presented in a winter school in Pushchino (near Moscow) and later published as a chapter in book edited by Alexander Levich. Published in: A.P. Levich [ed.] 1995. On the way to understanding the time phenomenon: The constructions of time in natural science. Part 1. Interdisciplinary time studies. World Scientific, Singapore. New Jersey. SummaryAccording to Meyen's concept, time is the variability (change) of real objects. Each object has its individual time, and a class of similar objects has a generalized time or time archetype. Typological and historical approaches are compatible if archetypes for morpho-processes are considered. Time in physics can be interpreted as a time archetype for all material objects. Meyen proposed the following principles of historic reconstructions on the base of his concept of time: typological extrapolation, process extrapolation, merono-taxonomic unconformity and multiple hypothesis testing. Further development of this concept is undertaken. It is shown that 1) object boundaries in space and time are based on archetypes, 2) object interactions affect their time by choosing development trajectories, 3) typological time is multidimensional, and each dimension characterizes the change of some particular part of the system. The principal components' analysis of multidimensional time allows one to distinguish between process and time, to define simultaneousness, and to formulate laws of system dynamics.IntroductionDifferent notions of time have developed in different natural sciences. Newtonian physics deals with absolute time and space considered as a world frame. In biology and geology time is build up of qualitatively different periods, such as eras or periods in geological history, seasons of year, stages of animal development. These periods correspond to certain changes of real objects and cannot be considered as absolute notions. For example, each geological period has its characteristic flora and fauna, each season is characterized by phenological phases of plants, and each development stage of an animal has specific morphological characters. Therefore, time is considered not as a world frame but as its canvas. It is not a background of object change but the change itself.Theoretical base of this point of view on the nature of time was developed by deceased Russian paleobotanist Sergei Meyen (1982, 1983, 1984). Meyen in his routine scientific work always had to reconstruct processes which had place in ancient times, for example, processes of plant development and evolution, processes of sedimentation and transformation of geological structures. The analysis of the essence of time was one of his most important objectives. His concept of time was based on Russian methodological traditions represented in publications of Vernadski (1975), Beklemishev (1969, 1970), Liubishchev (1982), Urmantsev (1971), and others. The conceptMeyen's concept of time can be best characterized by his own words: "From the observer's point of view, time is the variability of each object (individual) in the environment. This variability in some cases is receipted directly: different states of the same individual are projected by observer on his own variability, which can be called as 'psychological time' (or 'observer's time'). In other cases an apparatus replaces an observer. Observer considers his own variability as something a-priory defined. An idea of psychological time can be corrected by integration of data, reported by different observers (because observations should be repeatable). From this point of view, psychological time of a 'generalized observer' is a-posteriori defined. We have to consider general features in variability of some set of individuals because we are interested in receiving information concerning not individuals but classes of individuals (taxons). In result we receive the time of object class, i.e. an order of variability peculiar to each individual from the class, and receipted by some generalized observer. The lower is the taxon rank, the more complex and more detailed will be the corresponding time class. When extending taxon boundaries the similarity of its individuals decreases (an archetype of the taxon becomes more poor) and the corresponding class of time becomes less specific. The biggest taxon comprises all material objects in the world. The only common feature of all these objects is their materiality. Then material points will replace archetypes and variability will degenerate in its content and will become a generalized variability - an absolute time, designated by symbol t." (Meyen 1982, p.365-366, my translation from Russian).The following ideas can be emphasized in this fragment:
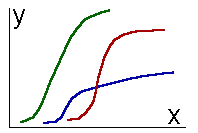
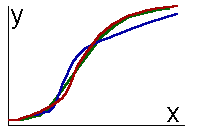
To reveal an archetype, it is necessary to study object structure: to fraction it into parts and study individual parts as well as links between them. The next step is the classification of parts according to their internal structure and their links. Meyen (1977) proposed a new term 'meron' for a class of similar parts. Therefore, meronomy is the classification of object parts. One meron embraces similar parts not only of one object but of many objects. Objects are considered as similar if they have common merons (their parts belong to the same meron). An object part can be considered as a trait or character if it is identified as a meron. In the first approximation, an archetype is a set of traits which are common to all representatives of the taxon (below it will be shown that this approach to the definition of an archetype is oversimplified). Typology is a study of object forms, but time destroys the form because the form changes. Thus, historic approach in science (especially in biology) was always considered as an alternative to typology. Meyen (1982) proposed a general idea that typological and historical approaches are compatible if the form of an object is considered in dynamic aspect. Then the form is not destroyed, but created by time. Of course, this idea is not absolutely new. For example, Beklemishev (1969, 1970) proposed to classify not instantaneous object states, but morpho-processes. Biological classification deals not with definitive stages of organisms but with their life cycles (definitive stages are used only if the life cycle is unknown). Temporal structure of an organism is not less important than its spatial structure. Meyen's merit is that he generalized this idea and formulated it as a methodological principle. He developed a system of notions for description of temporal structure of objects. For example, the notion 'meron' was applied to the life cycle of organisms: stages of development can be classified within one life cycle as well as in life cycles of some taxon. The class of similar life stages is a meron, which is considered as a 'generalized life stage'. Meyen called his concept of time as 'typological' because it is based on typological approach to the study of individual and group variability of morpho-processes. Principles of historic reconstructionMeyen (1984) applied the typological concept of time to develop principles of historic reconstruction (particularly in geology and paleontology). Any reconstruction is an extrapolation of patterns found in one kind of objects and phenomenons from other similar kind of objects and phenomenons. Meyen indicated that extrapolation was possible only on the base of typological analysis. He distinguished 2 kinds of extrapolations: taxonomic and meronomic. The first is applied to objects within one taxon, while the second is applied to object parts within one meron. "After entomologists had found giant chromosomes in salivary glands of several Drosophila species, they claimed that all Drosophila species have such chromosomes in salivary glands. This is a taxonomic reconstruction. We know that tracheids of coniferous trees have bordered pores. This feature is believed to be peculiar to all tracheids in the tree bole, although only a few preparations from different parts of the bole were studied. This is a meronomic extrapolation" (Meyen 1984, p.9, my translation).Meyen considered the principle of typological (taxonomic + meronomic) extrapolation as the first and most important principle of historic reconstruction. It can be used for creating hypothesis concerning fossil objects. To identify an object (for example, a fossil animal) means to find its taxon. One can extrapolate features of identified object from features of other objects of the same taxon. In particular, features of contemporary animals and plants can be used for reconstruction of missing parts of fossils. Therefore the principle of actualism appeared to be a particular case of the principle of typological extrapolation. This principle can be used for reconstruction of individual time of objects. For example, the dynamics of tree growth is saved in the pattern of annual rings, which is a record of previous states of this tree. A structural part of the tree (annual ring) belongs to the same meron as a temporal part of the life cycle (previous tree state). A meronomic extrapolation takes place here. At the same time it is linked with taxonomic extrapolation, because it is assumed that in this tree annual rings represent tree growth in the same way as in other trees, which were studied previously. Objects which keep records of their previous states were called tempofixators (Meyen 1983). Tempofixation is one of most important features of human brain, it is the base for psychological time. Sedimentation rocks containing remains of fossil animals and plants are tempofixators as well. Some objects can separate parts of the body which keep the record of an object state. For example, insects and other arthropods molt and leave their chitin exuvium which keeps characters of the previous development stage. This kind of objects were called as temposeparants (Meyen 1983). Other objects which don't keep record of their previous states, were called as tempodesinents. The second principle is a principle of process reconstruction (or Bergson's principle). It means the possibility to reconstruct a continuous process on the base of discrete phases of variability. Observations are always represented by a terminate set of descriptions which can be associated with pictures in the motion-picture film. If the order of pictures is known and object deformation between them is small, then there are no problems with reconstruction of the process. But in paleontology process reconstruction is linked with great difficulties. First, it is often impossible to find the natural order of 'pictures'. For example, in evolution reconstruction it is possible to compare the age of fossil objects only if they are found in the same place (the order of sedimentation layers corresponds to the time axis). But in majority of cases it is necessary to build an evolutionary sequence out of specimens found in different places. The result of reconstruction is uncertain in this case . Usually biologists try to compile a morphological sequence and then interpret it as an evolutionary sequence. Second, paleontologist cannot be sure that 'pictures' are really taken from the same 'motion picture film'. Morphological analysis tells only that two compared pictures were taken from similar (or not similar) 'films', and there is no guarantee that they belong to the same 'film'. Thus only the time archetype can be reconstructed, but not individual time. Third, individual 'pictures' are often incomplete and therefore missing parts should be reconstructed. Therefore, application of Bergson's principle has many obstacles. Meyen (1984) emphasized the link of this principle with the principle of historic reconstruction because compiling of pictures into a motion-picture film is based on typological extrapolation. To my mind, process reconstruction is just a particular case of typological extrapolation because the reconstruction of intermediate states is a meronomic extrapolation within the individual time of an object. The third principle was called a merono-taxonomic unconformity. It means that internal polymorphism of taxons makes all extrapolations uncertain to some extent. Hence an archetype cannot be considered as a set of common features of all objects in a taxon because it may be impossible to find even one trait common to all objects in this taxon. Nevertheless such a taxon is natural if it has its specific appearance and is well distinguished from other taxons. An archetype of such a taxon is not a definite form but a law of polymorphism. This law can be formulated 1) by frequency distributions of traits and 2) by sequences of form modification. Meyen (1974) analyzed plant leaf variability and revealed a set of shape transformations by which one can receive a complete potential diversity of leaf shapes. Only a fragment of this diversity has been realized in the process of evolution. Meyen associated an application of the same transformation to different initial forms with theme development in a musical composition. Therefore, he named this phenomenon as 'refrain'. Refrain is an archetype character but much more complex than an ordinary morphological trait. The fourth is the principle of multiple hypothesis (or the principle of Chamberlain), according to which it is important to consider not one but a set of alternative models of reality. This set should be as wide as possible. This approach allows one to avoid subjective preference of some particular model and gives an opportunity to optimize model selection. Several additional principles were considered for identification and dating of fossil objects. But these are more specific and will lead us away from the theme. Problems of the typological concept of timeIn this part of the article I discuss several problems of the typological concept of time which were not considered by Meyen. These problems appeared when I tried to apply Meyen's concept to living organisms. I hope, this discussion clarifies some hidden difficulties of the theory and indicates approaches for mathematical modeling of time archetypes.Object boundaries in space and time.The first problem is the selection of objects and revealing their bounds in space and time. Meyen ignored this problem considering objects as a given reality. But this problem is very important for the whole concept. For example, the following questions appear: 1) what happens with individual time at the moment of vegetative reproduction, 2) what happens with individual time of gametes when they merge, 3) is it possible that the object disappears but later appears again, or such a 'resurrection' is logically prohibited?To my mind, the world is not composed of objects. Objects are detached from environment by observer's consciousness. This idea was expressed by Martynenko and Chebanov (1988) in respect to texts. Therefore, the distinction between taxons and merons is relational: if the whole world is considered as an object, then taxons will become merons. Our ability to detach objects is based on a-priory notion about object archetype. For example, the space boundaries of vertebrate animals are considered to coincide with their skin surface. But in mollusks the shell is considered as a part of an organism. Colonial hydroids have no distinct boundaries between organisms, which can be separated only by careful morphological analysis. Therefore, we see objects only because we know their archetypes. A logical circle can be noticed here because, on one hand, objects are detached using archetypes and on the other hand, archetypes are revealed on the base of object analysis. Such logical circles are common to typological approach. Another example is that taxon bounds are established on the base of an archetype, and an archetype can be found by examining objects within taxon boundaries. In all cases entering a logical circle is possible only through spiral movement, in the same way as a screw cannot be directly inserted into a nut but can be screwed in. The following example will clarify what such a spiral movement means. When a man starts examining the object he detaches an object from the substrate and obtains some primary ideas about object bounds. Comparing this object with other objects he can reveal some preliminary archetype. On the base of this archetype the boundaries of the object can be reconsidered. Then new object boundaries are used for reconsideration of the archetype and so on. It may happen that initially detached object is a part of a bigger whole system. The only criterion of truth is the informational value of an archetype (the problem of the value of information is discussed elsewhere, see Sharov 1991). Informational criterion of archetype naturalness was first proposed by Liubishchev (1982). Therefore object boundaries are correct if they allow to receive the most informationally valuable archetype. Object bounds should be considered not only in space, but in time as well. They are set according to the archetype of individual time. Temporal boundaries of multicellular organism are zygote formation (beginning) and death (end). Usually these bounds are conventional to some extent. For example, the death of a mammal can be dated either by heart stop or by irreversible changes of the brain. The beginning of life is sometimes considered not from the zygote formation but from the birth. Objects form a hierarchy because their parts can be considered as individual objects. Different hierarchical levels have specific rules for setting spatial and temporal bounds. For example, these rules are different for cells, organisms, populations. Hierarchical structure of objects is transferred to the structure of time, making it hierarchical (Levich 1986). Hierarchical structure of objects and their times allows one to solve some difficult problems in the typological concept of time. For example, vegitatively reproducing organisms can be considered as objects of the same hierarchical level, while the clone as a whole is an object of the next level. The clone is an object because it has specific processes such as the process of degeneration. If necessary, a more detailed hierarchy can be introduced. Two bacterial cells descending from one parental cell can be considered as an object "two cells" which is associated with the higher level of hierarchy as compared with individual cells. Therefore, parental cell and its individual time do not disappear in the process of division, it is transformed into the object "two cells". The object "two cells" can be considered until it is something more than just the sum of two cells, i.e. there are some relations between these cells (for example, they can form a chain). Now the transformation of time in the process of predator-prey interaction will be considered. At the moment of predator-prey contact a new object (predator + prey) appears at the next level of hierarchy. An individual time of this object starts from the moment of predator-prey contact (visual or material) and continue during processes of consumption and digestion. Predator individual time goes simultaneously - it is linked with processes of development, growth, maturation etc. After the end of digestion the time of "predator+prey" is reduced to the time of predator itself, and there is no more sense to consider them separately. Individual time of the prey terminates during its interaction with the predator because all its specific processes stop. Interacting objects always can be considered as a whole system with its specific time which cannot be reduced to individual times of interacting objects. Human society is a good example of such a system with specific laws of development. Continuity in space-time is an important feature of each object. Unconnected parts cannot interact, and therefore a discontinuous object is not whole. Time and system interactionsThe second problem is that in the frame of individual time it is difficult to consider interactions with other objects and with environment. Interactions change specific object processes, and therefore, they affect its individual time. We used to consider time as an independent variable, but here it appears to be dependent on external effects. Individual time can completely disappear due to some interactions, as an individual time of prey does in the process of interaction with a predator.Interactions are incompatible with a simplified notion of individual time, considered as a trajectory in the phase space. It should be considered as a fibration of possible trajectories. Such kind of time I call as potential time and distinguish it from realized actual time - a definite trajectory in the phase space, selected in the course of object interactions with environment and other objects. Here appears the distinction between future and past time: future time is potential while past time is actual. This distinction should be considered only as local, because it is true only near the present time. Too far past is as indefinite as future. One of the most important kinds of interactions is synchronization when individual times become similar. In biology there are many examples of synchronization: diurnal and seasonal cycles of plants and animals are adjusted to corresponding rhythms of environment, seasonal cycles of parasites fit the seasonal cycles of their hosts. Multi-dimensional nature of timeHierarchical organization of objects imply complex relations between the time of the whole and times of parts. Each part has its individual time and a class of similar parts, i.e. meron, has its generalized time. The state of an organism can be characterized by a set of states of its merons. Therefore, the time of a taxon is always multidimensional: it is projected to the time of each meron as to the coordinate axis. Understanding of the multi-dimensional nature of time helps to develop methods for its analysis.Change of meron state can be interpreted as a process. The time of each object is characterized by the flow of all its processes. If all processes were strictly correlated then all of them could be easily represented by one generalized process and therefore, multidimensional space would be reduced to one dimension. But processes are usually discordant to some extent, and thus each dimension is independent. For example aging consist of several processes, and some of them can go faster or slower than usual (Dubina and Orlov 1987). This idea of multidimensional time differs from that proposed by Levich (1986). He links each time coordinate with specific level of hierarchy of object structure, while I consider dimensions to be linked with different merons on the same level of hierarchy. If the time of a taxon has too many dimensions, then it is difficult to characterize the laws of object dynamics. Thus, the problem to reduce the number of dimensions may arise. We shall assume that the state of each meron can be described by a real number (otherwise the problem cannot be solved using already developed algorithms). Then the number of dimensions can be reduced by the method of principal components. This method reveals generalized processes in object dynamics. Voitenko (1987) used it for the analysis of human aging. He revealed two main components of aging: orthogenic and pathogenic. The first corresponds to the normal type of aging without any kind of pathology, while the second is the development of diseases. Projection of individual time on principal components allows to predict all the processes with acceptable accuracy. The number of principal components is less than the number of processes, and therefore, the number of time dimensions is reduced. Physical time can be interpreted as a principal component of the time of all material objects - it is close to Meyen's (1982) point of view. Principal components are important for understanding the difference between the process and the time. The time of a taxon is represented by principal components and therefore, it is not equivalent to processes (times of merons). Each process can be plotted against the time of the taxon using projection to the subspace formed by principal components. An example is plotting of movement against physical time. Two states are considered as simultaneous if their projections on principal components coincide. An idea of reduction of time dimensions should be developed into a statistical procedure. Now I don't have such a universal procedure, but here are some ideas, how it should be organized. The major difficulty is to choose appropriate scales to measure advancement of each process. These scales should allow to converge together individual times and linearize them (linearization is necessary because the procedure of principal components is linear in its base). These scales will define a special geometry of the phase space. As an example, we shall consider trajectories of organism growth (fig.1). First, these curves should be superposed using simple operations such as moving and shrinking-stretching. The result is shown in fig.2. It is important to minimize the number of operations used. Otherwise it will be too much degrees of freedom and an archetype can be lost by inadequate convergence of trajectories.
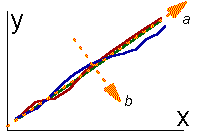
The second step is to linearize trajectories using nonlinear scales (fig.3). To describe the growth of living organisms Backman (1943) proposed to use logarithmic scale for calendar age (abscissa axis) and to modify the ordinate axis in accordance with the inverse function of normal probability distribution. These scales are shown in fig.3. When trajectories are straightened then the standard procedure of principal components can be applied. In fig.3 the principal component (a) corresponds to the Backman's law of biological growth , while the component (b) corresponds to the deviation from this law. This example demonstrates how the typological concept of time can be applied for description of generalized laws of object dynamics. Typological concept of time is most suitable for description of complex and organized systems. The criterion of suitability may be the degree of reduction of time dimensions when using principal component analysis. In simple systems there are only few processes, and therefore, there is no need to reduce dimensions. In weakly-organized systems processes do not interact, and thus, the number of dimensions cannot be reduced. In such systems each process can be analyzed individually. Only in complex and organized systems the number of time dimensions can be reduced considerably.
Acknowledgements:I wish to thank A.P.Levich, G.E.Mikhailovski, A.M.Maurin and V.M.Sarychev and other participants of Moscow Seminar for the Study of Time in Natural Sciences, for discussion of this paper.ReferencesBackman G. 1943. Wachstum und organische Zeit. Leipzig.Beklemishev V.N. 1969. Principles of comparative anatomy of invertebrates. Univ. of Chicago Press, Chicago. Beklemishev V.N. 1970. Principles of comparative parasitology. Nauka, Moscow (in Russian). Bergson H. 1911. Creative evolution. Holt & Co., New York. Dubina T.L., Orlov M.M. 1987. Patterns of age-specific changes in physiological characteristics and aging types. In: "Itogi Nauki i Techniki" (Review of Science and Technique), ser. General problems of biology, Moscow, 6:6-63 (in Russian). Levich A.P. 1986. Thesises on the time of biological systems. In: "Ekologicheski Prognoz" (Ecological Prediction), Moscow Univ. Press, Moscow (in Russian). Liubishchev A.A. 1982. Problems of form, systematics and evolution of organisms. Nauka, Moscow (in Russian). Martynenko G.Ya. and Chebanov S.V. 1988. Classification problems of stylemerty. Acta et commentationes Universitatis Tartuensis. 827: 119-136 (in Russian). Meyen S.V. 1974. About correlation between nomogenetic and tychogenetic aspects of evolution. Zhurnal obshchei biologii (Journal of general biology), 35: 353-364 (in Russian). Meyen S.V. 1977. Taxonomy and meronomy. In: Methodological problems in geological sciences. Kiev, p. 36-33 (in Russian). Meyen S.V. 1978. Principal aspects of organism typology. Zhurnal obshchei biologii (Journal of general biology), 39: 495-508 (in Russian). Meyen S.V. 1982. Methodology of the study of temporal relations in geology. In: Development of time concept in geology, Naukova Dumka, Kiev, p.361-381 (in Russian). Meyen S.V. 1983. The notion of time and typology of objects (in geology and biology, as an example). In: Dialectics in sciences about nature and man. Moscow, p. 311-317 (in Russian). Meyen S.V. 1984. The principles of historic reconstructions in biology. In: Systemness and Evolution. Moscow, p. 7-32 (in Russian). Meyen S.V. and Schreider J.A. 1976. Methodological aspects of classification theory. Voprosy filisofii (Problems of philosophy), N12: 67-79 (in Russian). Panova N.S. and Schreider J.A. 1975. Principle of duality in the theory of classification. Nauchno-tehnicheskaya informatsiya (Scientifical and Technical Information), ser.2, N10: 3-10 (in Russian). Sharov A.A. 1992. Biosemiotics: a functional-evolutionary approach to the analysis of the sense of information. In: The Semiotic Web 1991, T.A. Sebeok and J.Umiker-Sebeok (eds.). Mouton de Gruyter, Berlin, pp. 345-373. Urmantsev J.A. 1971. Specifics of spatial and temporal relation in living nature. In: Space. Time. Motion. Moscow, p. 215-241 (in Russian). Vernadski V.I. Reflections of naturalist. Space and time in non-living and living nature. Nauka, Moscow (in Russian). Voitenko V.P. 1987. Sexual differences in human aging and mortality. In: "Itogi Nauki i Techniki" (Review of Science and Technique), ser. General problems of biology, Moscow, 6:64-105 (in Russian).
Alexei Sharov |