BIOSEMIOTICS: FUNCTIONAL-EVOLUTIONARY APPROACH TO THE ANALYSIS OF THE SENSE OF INFORMATION
Alexei A. Sharov(Final version can be found in: T.A.Sebeok and J. Umiker-Sebeok (eds). Biosemiotics. The Semiotic Web 1991. Mouton de Gruyter, New York, 1992, pp. 345-373.)
Summary
The subject of biosemiotics, a new inter-discipline branch of science, is investigation of the biological nature of signs and semiotic base of biology. Information is considered as a micro-state of a system affecting the choice of system trajectories at bifurcation points. Sense of information has two components: meaning and value. Meaning is a set of bans and limitations set by information on the trajectories of system development and behavior, and value is measured by the contribution of information to the safety of self-maintenance and self-reproduction of the system. Meaning and value are considered at the material and ideal level. Sense evolution is characterized by its extension over time and space, and by comlication of its structure. This process went gradually from pre-biological systems till the man.Introduction
The problem of the sense of information has a great value for science and philosophy. In science it concerns mainly biology and the humanities. Biologists are interested in the sense of genetic information and signals involved in animal communication. Biological evolution is the development of the sense of genetic information. In the humanities there is a problem of the sense and its evolution in languages, arts, culture. Close relation between problems in biology and social science is not accidental. It is the consequence of the common semiotic nature of both branches of science. All sign systems must have the same general pattern of their structure and evolution. This hypothesis lies in the base of general semiotics. There are no sign systems functioning out of touch with living beings, and no living organisms which can function without some sign system (at least genetic code). This fact gives evidence of common roots of biology and semiotics, and indicates the necessity of the synthesis of these two sciences.Uexkll (1909) was the first, who analyzed problems on the crossroad of biology and semiotics. He considered that animals were dwelling not simply in environment, but in the sensible world, where each object had some meaning. This world was named 'Umwelt'. Stepanov (1971) introduced the term 'biosemiotics' for the branch of science that deals with sign systems of living beings. In this context biosemiotics appears to be an applied branch of semiotics which adopts semiotic concepts to biology. I think we need the mutual dialogue between biology and semiotics, though. So, we shall interpret the term 'biosemiotics' wider, not only as borrowing semiotic terms into biology but also as enriching semiotics with biological concepts, especially with functional-evolutionary approach.
Somebody may argue that such an approach already exists in semiotics, and there is no reason for borrowing it from biology. Morris (1971) distinguished three aspects of semiotics: syntactics, semantics and pragmatics. The first deals with inter-sign relations, the second - with the meaning of signs (i.e. with relations between signs and outer world), and the third concerns function and evolution of signs. Pragmatic problems are widely discussed in modern semiotics (Nauta 1972, Eco 1977, Perron and Collins 1989). But they concern only linguistic pragmatics that deals with perception and interpretation of signs of some natural language. Nevertheless signs continue their function after perception and interpretation - they control man behavior. But this process is not analyzed in the frame of linguistic pragmatics.
Semiotics has serious obstacles in biology, because its main terms such as sense, meaning, signifier, signified, and others being specified for subjective human experience, loss their clarity when applied to animals and plants. Deliberate limitation of semiotic subject to human sign systems (Eco 1977) is in fact the capitulation in front of this problem. Many scientists share the point that there must be common semiotic laws in human society and in animal world (Stepanov 1971, Sebeok 1972, 1976). These laws, if they exist, have to be formulated in non-anthropomorphic terms, based not on subjective experience but on observable events. Such non-anthropomorphic interpretation of semiotic terms at first may seem to be strange and unusual, but I see no other way of solving the problem. It is impossible to invent a new language for biosemiotics - it would be ambiguous and inconvenient.
It is important to include physiological, ethological and evolutionary aspects of sign functioning into pragmatics. And at this point the application of biological approaches will become inevitable.
In the field of philosophy biosemiotical problems are important for understanding the essence of ideality. We always think about ideal world using anthropomorphous terms. These terms can be applied only to human beings, and therefore humanity seems to be something exceptional for its ability to enter the ideal world. As a result a man estranges himself from nature and this may be considered as one of the causes of ecological crisis. The unity of man and his environment can be achieved only by extension of notions, concerning ideality, over the bounds of anthropomorphism. After that we shall be able to find senses and values in our environment and to reconsider ethical norms of relations between man and nature.
1. Essence of information in pragmatic aspect
Analysis of the problem must begin with the definition of information. All existing theories of information specify its different properties (quantity, value, truth), but they don't give general definition. The theory of transmitting and coding of information (Shannon and Weaver 1949) does not touch the essence of the subject, because information does not function in the process of transmission and coding. It can be compared with sending of any thing by mail: the essence (purpose and function) of the thing has no importance for the post. The only important character is the weight of a parcel which has no more relation to the essence of the thing than entropy to the sense of information.In the theory of Kharkevich (1960) value of information is determined by the increase of probability of achieving some purpose. But the mechanism of this process is ignored, and therefore the notion of information is not discussed. Obviously, not only information can increase the probability of achieving the purpose, but resources and energy as well.
Semantic theory of information (Bar-Hillel 1964) is devoted to the problem of truth. But this approach is not universal. For example genetic information is neither true nor false.
Two forms of information will be distinguished: potential and actual. Potential information does not function and is separated from the receiver. Actual information is either initially bound with the system in which it functions, or percepted by such a system and controls its activity (it may be not only physical activity but mental activity as well). The essence of information is expressed only in its active form.
Conditions for the function of actual information are the following:
- the system with actual information must have spontaneous activity;
- information characters of the system must be incommensurably small as compared with structure characters;
- stable trajectories of system dynamics must have unstable bifurcation points;
- information characters must influence the choice of system trajectories at bifurcation points;
- bifurcation points must be protected from outer noise.
The question about the origin and sources of spontaneous system activity is not essential for us. We admit the existence of such activity as an axiom. This activity can be described using the Petri net theory (Peterson 1981, Sharov and Kull 1990). But mathematical modelling is behind the point in this work.
Informational characters and structure characters must belong to different hierarchical levels, the scale of the former being much less than the scale of the latter, because an information process is always associated with great response to a small signal (Ackoff and Emery 1972). For example, in biology nucleotide sequence of DNA is an informational character and the body shape is a structure character. Characters of animal mental state pertain to information as well. It will be enough to consider only two hierarchical levels of characters: macro- and micro-characters. They characterize the macro- and the micro-state of the system, correspondingly.
Differentiation of scales of macro- and micro-characters leads to their relative independence. Thus we can consider them as two worlds: material and ideal. Material world is independent because its dynamics can be predicted along all stable trajectories. Ideal world can interfere in this process only at unstable bifurcation points. That is why such points are essential for the function of actual information. The more is the number of unstable points the more matter is 'spiritualized'. It becomes clear why physics predicts the movement of a stone perfectly but cannot predict the movement of a man.
Ideal world initially is not independent because it can be affected by material world at any time and at any extent. But evolution leads to the formation of autonomous regions in ideal world isolated from outer macro- and micro-characters. This question will be discussed in details later.
It is important to emphasize that ideal world is not less objective than the material one. It is not a product of imagination but a set of real system characters that can be examined either directly as in the case of genetic program, or indirectly as in the case of mental state (behavioristic methods in ethology).
According to the 4-th condition, information chooses the direction of system development at bifurcation points. It can be compared with branching railways, where point switches are controlled by information. Noise-protection of bifurcation points is necessary for the safety of information processes. Information must act specifically and therefore the spectrum of percepted inputs has to be very narrow.
So, I define actual information as a micro-state which controls the choice of system trajectories at bifurcation points.
Enumerated conditions of information functioning indicate that not every system is able to use some certain information. There must be close linkage between information and the structure of the system in which it functions. This statement contradicts with the common notion about information universality, that follows from the Church's thesis, that any calculation (in philosophic interpretation - any activity) can be executed by the universal Turing's machine. But this machine is a very simplified model that cannot represent specificity of information. Any material realization of the model will be not universal. It will work only if information is represented in appropriate form (I mean the format of the tape, configuration of signs etc.). Thus, in contrast to the thesis of information universality, I put forward the thesis of its specificity. Information functions only in systems that are fit to it. If there is lack of fitness, it will function only partially and not always adequate.
Living organisms are fitted to some definite genetic information. Gene expression is known to be regulated by the cell cytoplasm (Raff and Kaufman 1983) and thus cytoplasm is responsible for the specificity of genetic information. It is obvious that DNA of a man will not work in the cell of amoeba. Monophagy among viruses is the consequence of specificity of their genetic information because it functions only in the cells of certain host-species. Far hybridization of living organisms rarely gives good results because of the specificity of genetic information.
The role of genes in embryonic development can be explained using the idea of epigenetic landscape (Waddington 1968). Further we shall call it as epigenetic field because this term we consider to be more precise. Epigenetic field is a vector field in the space of states of an organism that determines its development. Stable trajectories were named creods. Genes determine the choice of developmental trajectories at branching points. But the set of creods exists independently from genes. Thus the statement that genes (in the fixed environment) completely determine organism characters is not true. Genes only select some variant from the given variety of potential forms. The dogma about exclusively genetic nature of heredity leads to a logical contradiction (Igamberdiev 1986, Sharov 1988) because semantics of any formal language cannot be set inside of the same language; it must be determined in a meta-language. In the morphogenesis of an organism the epigenetic field plays the role of a meta-language. Thus heredity cannot be reduced to the gene transfer from parents to their offspring. It is necessary to provide the descendant with a system capable to realize genetic information in morphological characters. An ovocyte is such a system. Therefore heredity has two unseparable levels: genetic and epigenetic.
Similar concept was developed by Zemek et al. (1985) who considered several levels of information transfer from parents to offspring: molecular, cellular, somatic and behavioral. But they did not indicate the difference between them: molecular and behavioral levels are characterized by the transition of information, while a system for realization of this information is transferred and specific epigenetic field is formed on cellular and somatic levels.
Now potential information can be defined more exactly. It does not function and thus enumerated conditions has no relation to it. Potential information for a system A is represented by its macro-characters or some characters of another system B, capable to affect actual information in the system A.
2. Meaning and value as two aspects of the sense of information
Our task is to give a general definition of sense, which can be applied not only to human languages but to animal communication systems and to genetic information. It's really difficult to suggest what is the sense of a gene: m-RNA, primary protein structure, its terminate structure, function or morphological character, or the corresponding character of an offspring that has inherited this gene.According to traditional semiotics, the sense of a sign is its mental content that is opposed to the meaning - a thing (or class of things) that corresponds to the sign (Frege 1892). But I think this interpretation of sense to be very narrow. Some obsolete slogan (for example, in Russia it is 'Forward to the victory of communism!') has no sense in spite of it has a certain mental content. What is lack in such a slogan is the value. Therefore I step aside from usual semiotic terminology. What Frege named as sense I shall call an ideal meaning and oppose it to material meaning. As to sense it is composed of the meaning (material and ideal) and the value.
Potential information when considered alone has no sense. But in connection with the receiver it will posess the sense, determined by its contribution to the actual information of this receiver.
2.1. Meaning
I define the meaning of actual information as a modification of probabilities of potential trajectories of a system under the informational control. It is reduced to a set of restrictions and bans of some ways of system development. Information has the meaning if there is at least one bifurcation point where it affects the choice of development or behavior trajectory.If the system has a fixed trajectory of development, then there is lack of meaning of any information. For example, any information about outer world has no meaning for a man convicted to a life imprisonment (in our theoretical world we shall exclude the opportunity of escape). Thus the punishment by deprivation of freedom has among others a semiotic aspect as a deprivation of meaning of information.
In the most simple case information affects the trajectory of the macro-state of the system. The field of macro-states will be called a macro-field. An epigenetic field is its particular case. The meaning of information in a macro-field we shall call a material meaning. Ideal meaning will be defined later on.
The meaning of information can be stratified into a time-series (fig.1). Let's admit that information prohibits the trajectory 'b' at the first bifurcation point and the trajectory 'd' at the second bifurcation point. Then its meaning is the state 'a' in the time-layer I, and the state 'c' in the time-layer II. In living beings the meaning of genetic information is not only the definitive stage of development, but the whole onthogenesis as well. Each stage of development is a separate layer of meaning. The meaning of genetic information extends to the progeny of an organism and has there additional layers.
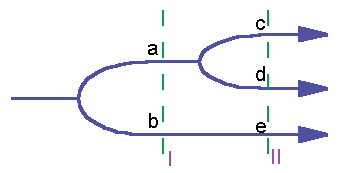 Fig. 1. Stratification of information
meaning in the macro-field (solid lines = stable trajectories; dashed
lines = time strata).
Fig. 1. Stratification of information
meaning in the macro-field (solid lines = stable trajectories; dashed
lines = time strata).
Now we shall complicate the model and define the meaning of a single micro-character of a system as its contribution to the meaning of the whole actual information. Micro-characters can interact in their effect on system dynamics. Multi-variate statistical methods can be used for separation of information meaning into components, corresponding to individual micro-characters and their interactions. This procedure is usual in traditional genetics aimed at the analysis of correspondence between genes and phenotypic characters.
An interesting type of interaction appears if some micro- character A gives the meaning to another micro-character B. For example, genes of insect wing venation display their meaning only if the wings are perfectly developed. The gene that prevents the development of wings in Drosophila fly deprives these genes of their meaning. It can be clarified using fig.1. Let's admit that trajectory 'a' corresponds to the presence of wings and trajectory 'b' - to their absence. The first bifurcation point is controlled by the gene of winglessness, and the second - by the gene determining the type of venation. The latter gene has its meaning only if the trajectory 'a' has been chosen.
A micro-character which gives the meaning to another micro-character we shall call a meta-information. This term was introduced by Schreider (1974) for preliminary knowledge necessary for adequate perception of information. Meta-information is a particular type of information which prepare trajectory bifurcations that will be controlled by other information. Genetic information is functionally heterogeneous because genes plays not only informational but also meta- informational role.
Meta-information geometrically can be represented in two equivalent ways: 1) as a choice of the trajectory that leads into the region of the macro-field where some other micro-character has its meaning, and 2) as a deformation of the macro-field. The first way that has been described previously is more convenient for theoretical analysis because all bifurcation points are considered to be fixed. But the second way is better for interpretation, because the appearance of the meaning of some micro-character is associated with the formation of the corresponding bifurcation point.
Now we shall consider another sort of meaning - ideal meaning. Let's admit that a system has an autonomous subset of micro-characters that is independent from other characters. The set of all its states is a field, that will be called a micro-field, because at any point there is a single vector of its change. Other micro-characters can affect the trajectory in the micro-field only at unstable bifurcation points. So the ideal meaning of a micro-character is its effect on the choice of trajectories in the micro-field. Consciousness is an example of a micro-field in which trajectories of thought depend on information percepted. In this case the micro-field can be interpreted as a semantic field.
Ideal meaning is peculiar only to those micro-characters that are not included into the micro-field, because otherwise the choice of trajectories will be not free - it will be determined inside of the micro-field. Therefore ideal meaning demands differentiation between micro-characters. But this differentiation is only functional and is not so fundamental as between macro- and micro-characters.
For example, the process of reading begins with formation of the visual image of the text in the mind. This image controls the trajectory of thought and leads to the understanding of the text. After the text has been understood the mental state of the reader is the ideal meaning of the visual image of the text. The material meaning will appear later when the man will change his behavior according to the information percepted. This example indicates that material meaning can arise only after ideal meaning. The delay may be so long that material meaning will remain unrealized till the death of a person. This delay appears due to memory. In primitive animals memory is poorly developed and so, there is no reason for analyzing ideal meaning, because it is immediately followed by correspondent material meaning. But ideal meaning is quite important for man, because it is the base of thinking.
The second example is a carcinogenic mutation in an organism. Biochemical and morphological characters are considered as micro- and macro-characters correspondingly. Then the ideal meaning of the mutation will be the reorganization of cell metabolism, and the material meaning - the cancer formation.
The third example concerns an automat that can project and produce things with defined features. Input features of a thing are initial micro-characters, their ideal meaning is a project, and material meaning is a thing being done. It must be emphasized that the function of the automat can be interrupted at the state of ideal meaning. In this case it will exist without material meaning.
Two informational states may have different ideal meanings but the same material meaning. For example, expressions 'the Morning Star' and 'the Evening Star' have different ideal meanings, which determine different algorithms for searching of the named object. But the material meaning is the same - it is the discovery of the planet Venus.
Material and ideal meanings can be compared with the 'meaning' and 'sense' by Frege (1892). The previous example borrowed from Frege indicates the close relation between these pairs of terms. Nevertheless our notions are dynamic and associated with system activity, while notions of Frege are static and abstracted from activity.
Meta-information can be considered not only for material but for ideal meaning as well. It will be a micro-character that supplies another micro-character with its ideal meaning. In human mind notions play the meta-informational role. Understanding of a notion is equivalent to getting into the region of semantic field, where information associated with this notion receives its meaning.
The region of semantic field where consciousness operates can be considered as assimilated thesaurus of a person. According to Schreider (1974), information can extend assimilated thesaurus and so it can serve as ideal meta-information.
2.2. Value
Value is the second component of the sense. In human society it is determined by usefulness of information in different spheres of activity: industry, culture etc. (Zunde 1982). Informational market is highly developed in postindustrial countries and the value of information has its money equivalent.The value of information is usually estimated according to the increase of probability to attain the purpose after getting the information (Kharkevich 1960). It may be positive and negative and it has no direct relation to the truth of information. So, in the theory of Kharkevich the problem of information value is reduced to the problem of purposes. But Kharkevich only postulates existence of purposes and does not consider axiological topics. This approach is not acceptable in biosemiotics: we cannot postulate what we want to prove.
According to Kharkevich, the purpose plays the role of meta-information because it gives the meaning to all additional information useful for its achievement. But as the purpose is a sort of information the question arises about the value of the purpose. However it is possible to construct a hierarchy of purposes, each of them being evaluated by the purpose of the next level. But the purpose of the highest level will have uncertain value. Therefore I think that value is a more fundamental notion than purpose, in spite of that in some cases purposes can be used for estimating values.
The purpose is always separated from the system because it perishes being combined with the system (if the purpose is attained there is no purpose more). But value is immanent to its bearer. It can be thought as a purpose being combined with a system. Value is 'entellehia' that means 'containing the purpose inside'.
The value unlike the purpose can be measured quantitatively by the coefficient of reproduction. This coefficient is less than one, if the system cannot reproduce itself, and may be more than one for self-reproducing systems. Thus, value exists due to processes of self-maintenance and self-reproduction. In self-reproducing systems the coefficient of reproduction can be made more than one if appropriate process rates are chosen (Sharov and Kull 1990). In the case of self-maintenance it is impossible. So, self-maintenance is qualitatively different from self-reproduction.
The expression that value is a purpose combined with a system, must be considered as a metaphor. There is no need for the notion 'purpose' while defining value. Value of the system is equivalent to its ability to maintain and (or) reproduce itself.
The value of information is measured by its contribution to the value of the system. If there are two systems with identical macro-states and different micro-states, then the system with greater coefficient of reproduction has more valuable actual information. In other words the value of information is measured by its ability to support survival and reproduction of the system, and therefore it is increased by natural selection. The value can be estimated not only for the whole micro-state of the system but for individual micro-characters and for their interactions as well according to their contribution to the value of the system.
The problem of identification of initial and terminate system states is essential for estimation of value, because we need a criterion for the life-cycle closure. The precision of identification must be chosen so that identified systems will have approximately equal coefficients of reproduction. Different sets of characters may be used for identification. The least set will contain all macro-characters essential for survival and reproduction of a system and all micro-characters determining the development of mentioned macro-characters. The value correspondent to this set of characters will be called a material value. Systems with equal material values may be compared by their ideal value - ability to keep and reproduce those micro-characters that have no effect on survival and reproduction in the chosen time-scale.
Now the model will be complicated by introducing several time-scales. Each scale has its specific values. For simplicity two scales will be considered: short and long. Here are several examples. Carcinogenic mutation in the cell has considerable short-scale value because cancer cells proliferate with greater rate than normal cells in an organism. Nevertheless, this mutation has no long-scale value because cancer leads to the death of an organism with all its cells. The second example is gigantism in animals: it has no long-scale value because it leads into the evolutionary dead-lock, but in favorable conditions its short-scale value may be rather high. Natural selection first operates on the short time-scale, and so systems with low short-scale value will be eliminated. But if systems have equal short-scale values then they will be selected according to their long-scale value. Such multi-level selection is called hierarchical adaptation (Conrad 1983), cascade selection (Lekevicius 1986), selection of taxons (Sharov 1979, 1984).
The next example is interesting because it concerns a very short time-scale as compared with the duration of a life cycle. Corresponding value consists in the stability of the present state of an organism and can be supported by adaptive behavior controlled by actual information. If this behavior ensures not only short-scale stability but also the closure of the life cycle with great coefficient of reproduction, then the correspondent actual information will have both short- and long-scale values.
We can consider such a short time-scale in which considered systems have equal material values. Then they can be compared by their ideal value. Two kinds of value (material and ideal) correspond to two forms of selection. Material selection is differential survival and reproduction and ideal selection is preference of some certain micro-states of a system. The more is the delay in the action of material selection the more clear will ideal selection be displayed. In particular, in human population material selection is extremely weak and therefore ideal selection is well developed. I dare say that ideal selection plays the main role in human evolution.
In primitive systems ideal selection is expressed in the nearing to the most stable micro-state. In more complex systems with a micro-field it will take the form of a free choice of states and trajectories in this field. All trajectories are primary equivalent, but ideal selection breaks this equivalence specifying preferable transitions between states. Selection is represented by a set of appraisals for states and trajectories in the micro-field. In this case the stable ideal meaning of information has the highest ideal value.
An example of ideal selection is the development of immune system in animals. Its state is characterized by a spectrum of antibodies in the blood. The set of states is a micro-field in which any state can be attained by small control inputs. Clone- selectional pattern of antibody production (Friemel and Brock, 1986) leads to the choice of a certain state of immune system, nevertheless all states are potentially equivalent. The chosen state is the most fit for protection of an organism. But this fitness is not obligatory and may be destroyed in the case of autoimmune reaction.
The state of the micro-field with great ideal value we shall call an ideal purpose, and its material meaning - a material purpose. Ideal purpose can be interpreted as an intention to attain material purpose. This intention may be either specified or abstract. In the first case the system instantly begins acting, and in the second case it begins formulating derivative purposes or passively waits for appropriate conditions for achieving the purpose.
Introduced definition of a purpose differs from the usual meaning of this term, because it corresponds not only to the terminate state of activity but to the whole process. In some cases there will be no terminate state at all. For example, such purposes as prosperity, honor, good have interminate material component. Previously we suggested to name them not 'purposes' but 'values' (Schreider and Sharov 1982). But nowadays I prefer to use the term 'purpose' in wide sense including the mentioned case because the ideal component of the purpose is achieved at the moment of decision-making.
Thus, the sense of information we consider to have two aspects: meaning and value. Meaning is a semantic characteristic of sense and value is a pragmatic characteristic. Information has sense only if it has both aspects. Actual information has its actual sense, and potential information has potential sense that is different for each receiver.
3. Evolution of the sense of information
In this section principal trends in evolution of information are to be discussed. The main task was to consider a continuous evolutionary line from primitive pre-biological systems till very complex processes in human society.Three main trends are seen in the evolution of the sense of information:
- the quantitative expansion in time and space,
- integration of its structure,
- development of potential information.
3.1. Expansion of sense
It's incredible that matter in the past had no information and no sense at all. Teilhard de Chardin (1965) was true considering non-living nature as a 'pre-life', containing ideality in its rudimental form. Nevertheless the problem of sense origin is not declined, because new senses arise in the the process of evolution.Sense expansion in time and space can be explained by natural selection. First we shall consider its time-extent. If information has value, i.e. it improves self-maintenance and self-reproduction of the system, then its preservation will increase the coefficient of reproduction. Thus, time-extension of the meaning of valuable information will have selective advantage. But information can possess some value only if it has considerable time-extension of the meaning. So the value and the meaning of information cannot arise separately. The only way of their becoming is iterative parallel development after their accidental appearance in some small threshold quantities.
An important stage of sense evolution was the appearance of informational self-reproduction, when a system became able to create its offspring with the same macro- and micro-state that it had had on some previous stage of development. Informational self-reproduction is the way of sense expansion over time because the sense of parental system is displayed in the long chain of offsprings.
The space coordinate is also important for self-maintenance and self-reproduction. The growth of the system allows it to intensify its interaction with environment. Informational control of remote objects stabilizes the system and ensures its safety.
Main mechanisms of space-expansion of the sense of information are the following: 1) movement, 2) polymerization and 3) communication.
A mobile system can affect remote things and fit itself into the situation. So an object in the environment appears to become the part of the system and can be used by it if necessary, as an animal uses its leg or tail. In this way external space turns into internal space of the system. It is functionally equivalent to the extension of body bounds.
When a system fits into the situation it creates the ideal image of its space. An image of any point in space is a subroutine of achieving this point. If there is a thing in the environment, that is valuable for the system, then its ideal image will be a subroutine of its using. So, fitting into the situation means creating a set of subroutines for movement and interaction with objects in the environment. This model can be considered as an explication of the notion 'Umwelt' by Von Uexkll (1909).
Fitting into the situation can be either passive or active. In the former case it is reduced to the internalization of the given world and in the latter case it means the modification of the world according to some program. Man differs from other animals primary by his active modification of the world he lives in. Satiation of space by artifacts helps him to create a great variety of niches and so improves his self-maintenance and self-reproduction.
Polymerisation is the duplication of information and its distribution between the copies of the systems. Informational self-reproduction is its particular form, characterized by complete division of systems. It allows an organism to distribute its information along large areas. But polymerization is not always followed by division of the system. The origin of multicellular organisms and colonial animals was based on such incomplete reproduction. In social systems, such as a colony of bees or ants, there are no morphological connections between its subunits but functional connections are highly developed.
Incomplete reproduction gives hierarchically structured systems and therefore the problem arises what is the correspondence between senses of information at different levels of hierarchy. Information of a super-system is a sum of informations of its subsystems, but the sense of information is not summed up because of interactions. Sociality extends the macro-field of subsystems and so the meaning of information becomes more complicated. Some micro-characters of an organism that has no meaning in solitary life will receive it in the society.
Social life affects not only the meaning but the value of information as well. For example, altruistic behavior becomes an alternative form of self-reproduction. According to the kin-selection theory (Hamilton 1964, Wilson 1980), it may appear to be more profitable for an organism in a colony to take care of its relatives than to increase its own fertility. The profit is measured by the number of genes transferred to the next generation. Relatives have similar genotypes and thus their reproduction fill compensate the reduced fertility of an organism with altruistic behavior.
Any society is heterogeneous and therefore conflicts may arise when different groups of organisms try to change the social structure in their own favor (Trivers, 1974). A conflict is a natural consequence of space extension of the sense of information, when different senses are being realized on the same matter. In primitive animals conflicts are not conscious and are resolved by the balance of natural selection. But man resolves conflicts on the base of his experience. There are 3 main ways for that: 1) to abandon the society, 2) to change the structure of society so that the conflict will be declined, and 3) to change one's own ideal values. So, in order to decline conflicts the society must give to its member: 1) the freedom to abandon the society, 2) the right to influence the direction of society evolution and 3) the freedom of conscience.
The second of these points demands the development of a system of limitations and bans for space-extension of the sense of individual information. One of such limitation mechanisms is property, which in primitive cases takes the form of territorialism. Property is a result of mutual commitment between society members about the division of 'spheres of interest'. A more comprehensive description of social patterns and evolution is given in the monograph of Boulding (1981). This book is discussed in details below.
Communication is the production and transmission of information not combined with system division. This mechanism of space expansion of the sence will be discussed later, because we need some additional termes that will be introduced in the next section.
3.2. Integration of sense structure
The structure of sense is a system of interactions between the senses of individual micro-characters and complexes of such characters. Apparently, in pre-biological systems the structure of sense was very simple with minimal interactions. All information was represented by a set of independent stable micro-characters, coding an algorithm of activity that is valuable in so far as the system using this algorithm still exists or (and) successfully reproduces itself.Variable micro-characters theoretically can have some value if they can change system behavior in accordance with environmental fluctuations. But it is almost incredible that some micro-character responses to the changes of environment and at the same time adequately affects the system behavior. It is more probable, that different variable micro-characters are responsible for indicating environmental changes and for control of system activity. But they must be connected by some stable micro-character. Therefore variable micro-characters taken alone have no value but their interaction with stable micro-characters that coordinates their work has considerable value. This interaction can provide self-regulation of the system based on the feed-back loop principle: the variable micro-character responses to the macro-state of the system, and its interaction with other micro-characters affects the further dynamics of the macro-state.
The state of regulated system is drawing to some attractor, in the simplest case - to a stable point. This attractor will be called a quasi-purpose which differs from the real purpose for it is determined by interaction between micro- and macro- characters, while the purpose is determined only by micro-characters. It's not correct to say that the system aspires to the quasi-purpose because the quasi-purpose is not distinguished on the ideal level.
A quasi-purpose must be more valuable than other micro-states because otherwise the system will fail in the struggle for existence. Thus, only those micro-states can become quasi-purposes, that have material value, i.e. they support self-maintenance and self-reproduction of the system.
Further development of interactions between micro-characters leads to the appearance of a micro-field. An informational mechanism, regulating system position in the micro-field will be called an operator. It creates its own ideal values and corresponding purposes. Ideal purpose is the ideal meaning of the operator, that can change in accordance with the macro-state of the system (including the state of environment).
If memory is absent, then an ideal meaning of operator depends on the current macro-state of the system only, and does not depend on previous experience. It can be changed only by natural selection. If the operator forms such ideal values, that in the greater time-scale increase the probability of system destruction, then it will have little material value and will be replaced by another operator in the course of natural selection. So ideal values in the short time-scale will be set in accordance with material values in the long-time scale.
A biological example is an instinct behavior in animals. It is inherited and evolves by natural selection. An animal acts in purposeful manner, but its purpose is not conscious. It knows what it is doing but does not know that it can be done in other way. Ideal purpose in this case is reduced to an algorithm of activity.
In the more complicated case, when memory is developed, ideal meaning of operator depends on previous experience of the system. Here a negative feed-back loop appears and therefore training becomes possible. Ideal purpose has complex structure: it consists 1) of a program of activity and 2) of an ideal image of the purpose (fig. 2). These components are formed by an operator in different micro-fields, one of which controls system activity and the other models its future macro-state. The appearance of modelling micro-field allows the system to reflect its purpose and therefore the purpose becomes conscious.
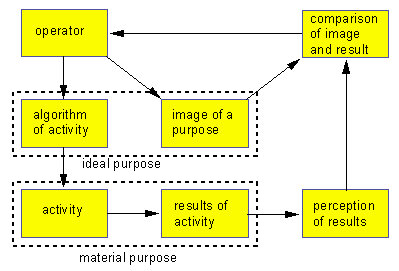 Fig. 2. Scheme of system training.
Fig. 2. Scheme of system training.
After the ideal purpose is formed it will control system activity and the result will be compared with the image. If there is no correspondence between the result and the image, then the material purpose is said to be not achieved. In this case the ideal value of the algorithm of activity will be diminished by the operator and it will be chosen less often later on. If material purpose is achieved the ideal value of the activity algorithm will be increased. In result ideal values will become adequate to the image of the purpose.
An example is conditioning in animals. If an animal has a conscious purpose to get food it forms and realizes some program of activity. If this program appears to be successful then it will be used more often.
In this model the image of the purpose cannot be modified by training. It is constant and may be replaced only by natural selection. For example if an animal has a purpose to get food it will touch the food placed in a trap and will die. Only those animals will survive that can distinguish between nature food and the food in the trap. But this concerns an abstract animal with only one purpose. Real animals have many purposes operating either in turn or simultaneously. In the latter case there must be some coordination between purposes, if they are incompatible. So, a hierarchical order in the set of purposes must be developed in the special micro-field. For example, the purpose of being ease is one of the strongest and in emergency it ceased realization of all other purposes. When an animal percepts the smell of iron it will not touch the food in the trap.
An operator that forms a conscious purpose is a stable micro-character in the majority of animals and is controlled by natural selection. Only man and may be some higher animals are able to choose it by their will. In this case a set of operators becomes a micro-field and a meta-operator is formed to control the choice of operators. It is also possible to consider meta-meta-operators and so on. The choice of operators is determined by their ideal value. But this value corresponds to the longer time-scale than values formed by these operators. For example, it takes little time to train how to do some certain thing, but it takes much longer time to learn how to choose what thing must be done.
It is obvious that the hierarchy of operators cannot be very high, because otherwise it cannot function effectively. The time required for training increases enormously with the level of hierarchy of operators. Therefore purposes and values of higher levels are selected for a very long time. Unless they are not selected on the base of experience their choice and keeping will be deliberate and not related to the basic material value - self-maintenance and self-reproduction. It explains the phenomenon of a free will. A man is strong-willed if he keeps ideal values of higher levels of hierarchy (these values are not always highly moral). The choice of these values is free because they are not controlled by selection for a long period of time, may be for the whole life. Therefore the will is always free.
3.3. Development of potential information
Potential information has two types of influence upon the micro-state of the system: functional and arbitrary. It affects the system functionally if the process of its assimilation is determined by physical properties of the carrier of potential information and receptive organs of the system. If the influence of potential information is realized by the choice of trajectories at bifurcation points of the micro-field, then it will be called arbitrary. Potential information that has only functional influence on the micro-state of the system I shall call a pseudo-sign. If both types of influence are present, it will be called a sign.Functional component is obligatory, because potential information first interacts with some sensitive organ that turns it into actual form. This process is always determined by the physical nature of the sign or pseudo-sign and therefore corresponds to the functional type of influence. Arbitrary influence appears later - after percepted information has started its control of the choice of trajectories in the micro- field. Thus, ideal meaning of a sign consists of two parts: functional and arbitrary. The first is determined by the physical properties of a sign and corresponds to the signifier by Saussure (1959), and the second corresponds to the signified. For example, the visual image of a road-sign in the consciousness of a driver is a signifier, and an algorithm of his activity, associated with this sign, is a signified. This association is conventional and corresponds to the road-rules.
Ideal meaning of a pseudo-sign does not split into signifier and signified. So, its effect on the system is not regulated at the ideal level. Therefore it is always unfavorable. Examples are alcohol and other poisons affecting nervous system of an animal, and some times affecting genetic system (chemical mutagens). Gamma-radiation also has mutagenic effect. In the course of evolution animals possessed protection mechanisms from natural pseudo-signs. Thus nearly all effective pseudo-signs are artifacts.
Signs can arise in evolution only if the correspondence between signifier and signified has the value. So, this correspondence is not arbitrary from the evolutionary point of view For example, a wolf percepting the smell of a rabbit enforces its searching activity. If its activity were always low the wolf would never find a rabbit. If it were always high then too much energy would be spent. So, it is advantageous for the wolf to combine perception of certain smell (signifier) with increasing of searching activity (signified).
The value of signs is an essential and sufficient condition for their origin in the course of natural selection. Primarily, I think, signs had their value only for receivers. There was no active production of signs but some natural things began to serve as signs. Such production was started only after signs had acquired their value for transmitters. A sign has value for a transmitter if its emission increases the coefficient of reproduction of the system. Particularly, in human society emission of texts is a source of welfare for an author and his family (fees, reputation, connections), and on the other hand, it gives the author an opportunity to spread his ideas - that is an analogy of self-reproduction.
A sign that has no value (no positive value) for the transmitter will be called a stimulus. For example, the smell of a rabbit is a stimulus for a wolf, because it would be better for the rabbit not to make smelling trails. Photoperiod is a stimulus for insects and plants to prepare for the winter.
A reverse situation may occur, when a sign has positive value for transmitter but negative value for receiver. Such a sign may be called an inverse sign. For example, females of some species of fire-flies imitate light signals of other species and so attract males and eat them. Another example is mimicry. Inverse sign is always an imitation of some other sign with positive value for the receiver.
A sign that has sense both for transmitter and for receiver will be called a proper sign. This sort of signs is the most common. Examples are sex pheromones of insects, acoustic communication of grasshoppers and birds, ritualized behavior of some birds and mammals, human speech.
Communication with proper signs exists only between close relatives: usually between members of one biological species. It can be explained by the similarity of macro- and micro-fields of related organisms. Therefore, information that has some sense (meaning and value) for one organism will have the same sense for another organism. The simplest but very effective type of communication is imitation that is used for synchronization of activity of several animals for attaining the common purpose. If a transmitter and receiver are sufficiently different, the message has different sense for them. An example is mating communication in animals. In this case precise coordination of senses is necessary that can be achieved only between closely related partners.
In hierarchically organized systems sign communication can function on several structural levels. In multicellular organisms cells exchange with chemical messages: hormones, mediators. Complex system of chemical communication exists on the subcellular level as well. Semiotics of internal communication is usually called endosemiotics (Deely 1982). Apparently, internal communication originated from external one in the process of development of a new level of organization.
The main trend in sign evolution is increasing flexibility of the link between the signifier and the signified (Sebeok 1976). This link is very stable in primitive organisms (pheromone communication in insects): it is inherited and changes only by natural selection. But higher animals and man have flexible link between the signifier and signified, formed on the base of private experience.
The highest level of communication is language. It becomes necessary when the number of signs increases sufficiently. The only way to keep the simplicity of the growing sign system is to use complex signs composed of simple subunits. It is important that the sense of a complex sign can be reconstructed out of senses of its parts. So, in language a sign has structural and sense organizations and a homomorphism between them.
In glossemantics these two organizations were called the plan of expression and the plan of content (Hjelmslev, 1961). The plan of content concerns the sense of signs and therefore it has two aspects: meaning and value.
Intensive sign exchange leads to the development of communicational Umwelt - a system of trust rates to different sources of information. It considerably improves the orientation of a system in its information environment. The sense of information becomes more and more dependent on the authorship, and anonymous signs lose their value. The rate of trust to the source of information depends on the correspondence between its signs and ideal values of the receiver. Therefore, any sign bears additional sense concerning evaluation of the source of information. This additional sense is connotation.
Communication leads to the fast distribution of ideal values and so enforces their development. Apparently, ideal values of higher levels of hierarchy cannot arise without communication.
Discussion
The branch of science named biosemiotics is only at the beginning of its development, and thus there are few pertaining references. Now I want to discuss briefly 3 concepts most related to biosemiotics: sociobiology (Wilson 1980), zoosemiotics (Sebeok 1972, 1976) and ecodynamics (Boulding 1981).Sociobiology applies functional-evolutionary approach for explanation of social behavior of animals and this draws it together with biosemiotics. Nevertheless it deals mainly with hereditary characters and does not touch general problems concerning the essence of information, sense and sign. Therefore, it is the least close to biosemiotics as compared with two other enumerated concepts.
Zoosemiotics of Sebeok is a wide-range classification approach towards general semiotics. His main merit is bold extension of semiotic notions over the limits of anthropo-semiotics. He proposed the classification of branches of general semiotics accordingly to the type of transmitter and receiver, the classification of signs by their material nature, function and mode of link between the signifier and the signified and the classification of communication channels. But his fidelity to semiotic traditions of Pierce and Morris prevents him from using functional-evolutionary approach.
The monograph of Boulding (1981) 'Ecodynamics: a new theory of social evolution' is the most interesting from biosemiotical point of view. He proposes the general concept of evolution as an expansion of systems over the set of 'niches' (generalization of ecological niches) and applies it to chemical reactions, biological species and human artifacts (machines, public organizations, languages etc.). New niches becomes occupied in result of 'mutations' (in broad sense).
Systems involved in evolutionary process are characterized by the 'phenotype' and the 'genotype'. The genotype or 'know-how' is an ability of a system to reproduce its own phenotype and phenotypes of other objects. The genotype of higher animals and man has two parts: 1) biogenetic part, coding morphological and physiological characters and 2) noogenetic part, coding development of neural system and behavior. Biological evolution up to man is characterized mainly by biogenetic change, but human evolution is mainly of noogenetic character. Noogenetic evolution differs from biogenetic one, first, by wide informational exchange, and second, by occupation of new niches by human artifacts. Ecodynamics is a considerable step forward as compared with sociobiology because it deals both with biogenetic and noogenetic changes.
The present article is very close to 'Ecodynamics' in its main principles. Nevertheless it is necessary to mark the difference and especially those points, where, as I think, I have advanced.
First, Boulding does not mention the semiotical base of biology. While Sebeok transmits notions from the humanities to biology, Boulding does the opposite thing, transmitting biological notions to social science. But in both cases there is a lack of feed-back loops and therefore both approaches are one-sided. In present work I endeavor to develop a mutual link between biology and semiotics. In particular, I consider that genetic information has meaning and value and therefore it has the sense. Coordination of ideal and material values in living systems is described. An attempt is made to explain the phenomenon of free will by hierarchical structure of human micro-state.
Second, Boulding eludes the discussion about mechanisms of information influence on the development and behavior of systems. In present model I suppose that information is represented by micro-characters of the system, capable to affect the choice of its trajectory at bifurcation points. The meaning of information is being realized in this manner either in the macro-field (generalization of epigenetic landscape) or in the micro-field (generalization of semantic field). Hence goes the double nature of heredity (genetic and epigenetic); it becomes clear why natural languages can exist without a meta-language, specifying the meaning for all signs.
Third, Boulding has no developed concept of value, and particularly he does not discuss the problem of information value. He only asserts that subjective human values play the same role as the natural selection in animals and plants - they support selective reproduction of certain systems. In present work the notion 'value' is used considerably wider. Subjective evaluation is only one specific sort of values - the ideal value. But besides there is material value expressed in the ability of systems to maintain and reproduce themselves. Material value is the base for development of ideal values, which are formed either by natural selection or by training.
In conclusion we shall discuss some consequences and applications of our theoretical model.
1. It is important to realize that informational processes and their evolution in non-living, living and conscious systems have the common nature and common laws. Therefore it is necessary first, to compile facts confirming these laws, and second, to develop the theory of global evolution. Apparently, natural selection will be the basic principle of such a theory (Csnyi 1982). It must be developed using generalized mathematical models, applicable to any natural and artificial systems. It is also necessary to develop a unified terminology in biology, semiotics, sociology and information theory. First steps have been done in the present work. The most essential notions are meaning and value as two sides of sense, and distinction between material and ideal components of meaning and value.
2. In the field of biology it is important to realize the semiotic nature of this science. The central biological notion - homology - has an informational base: homological organs are formed by similar genetic and epigenetic mechanisms. Therefore, homology has two aspects: genetic and epigenetic, that must be distinguished. Molecular biology deals mainly with genetic aspect of homology while traditional biology - with the complex interaction of both. Informational approach may be applied to the notion of progressive evolution. On the base of this work the following features of progress may be pointed out: expansion of sense in time and space, integration of sense structure and development of potential information.
3. In the field of ethology it is valuable to develop models for the structure and evolution of Umwelt in different groups of living organisms. The general theory of territorialism and competition may arise from these models.
4. Biosemiotic approach may be useful for the theory of biosociality. Necessary and sufficient conditions for establishment of social relations between organisms have to be revealed. Development of cooperation and communication can be described in terms of meaning and value of information. In particular, this approach can be applied to anthropogenesis and to the process of evolution of communication in human populations.
5. It is possible to develop a biosemiotical theory of property and a theory of conflicts. On this base a synthesis of ecology, economy and politology is possible. Many interesting ideas on this theme may be borrowed from Boulding (1981).
6. In the field of semiotics functional-evolutionary approach has to be applied. It allows to extend semiotic notions to the sphere of all information processes. The main conclusion of this work is that sense must be considered in its dynamic pattern. Semiodynamics is a new level of semiotics. In this work the model of sense realization by informational control of system dynamics in macro- and micro-field is presented. It can be used in computer models of artificial brain and expert systems.
7. Biosemiotic approach can be used in philosophy, especially in axiology. It clarifies such notions as ideality, purpose, value.
8. In ethics biosemiotics leads to semiocentrism, according to which progress consists in accumulation and complication of the sense of information. Therefore an imperative arises to keep all semiotic resources that have been accumulated in the biosphere of the Earth. All living organisms as carriers of sensible information have to be protected. Man has a special value because he has not only species-specific information but unique individual information as well. Favourable conditions must be created for intensive development of such information. Semiocentrism is able to normalize relations between man and nature, because unlike anthropocentrism it considers the man as a part of nature and does not oppose him to the nature (from the report of K.I.Yakimets, see Sharov 1990).
References
Ackoff, R.L. and Emery, F.E. (1972). On purposeful systems. Chicago, New York: Aldine-Atherton.Bar-Hillel, Y. (1964). Language and information. Jerusalem: Jersalem Acad Press.
Boulding, K.E. (1981). Ecodynamics: a new theory of social evolution. Beverly Hills, London: Sage publications.
Brennenstuhl, W. (1982). Control and ability: towards a biocbernetics of language. Amsterdam: Benjamins.
Conrad, M. (1983). Adaptability: the significance of variability from molecule to ecosystem. New York: Plenum Press.
Csnyi, V. (1982). General theory of evolution. Budapest: Publ. House of the Hung. Acad. Sci.
Deely, J. (1982). Introducing semiotic: Its history and doctrine. Bloomington: Indiana Univ. Press.
Eco, U. (1977). A theory of semiotics. London: Macmillan. Frege, G. (1892). Uber Sinn und Bedeutung. Z. fur Philosophie und Philosophische Kritik 100(1), 25-50.
Friemel, H. and Brock, J. (1986). Grundlagen der Immunologie. Berlin: Akad.-Verlag.
Hamilton, W.D. (1964). The genetical evolution of social behaviour. J. Theor. Biol. 7(1), 1-52.
Hjelmslev, L. (1961). Prolegomena to a theory of language. Madison: Univ. of Wisconsin Press.
Igamberdiev, A.U. (1986). Problems of description of epigenetic systems. Zhurnal obshchej biologii (Journal of general biology) 47(5), 592-600 (in Russian).
Kharkevich, A.A. (1960). On the value of information. In Problemy Kibernetiki (Problems of Cybernetics) 4, 54-58. Moskow, USSR: Nauka (in Russian).
Lekevicius, E. (1986). Outlines of the general adaptation theory. Vilnus: Mosklas Publishers (in Russian).
Morris, Ch. (1971). Writings on the general theory of signs. Hague: Mouton.
Nauta, D. (1972). The meaning of information. Hague: Mouton.
Perron, P. and Collins, F. (eds.) (1989). Paris school Semiotics. 1.Theory. Amsterdam (Philadelphia): J.Benjamins Publ. Co.
Peterson, J.L. (1981). Petri net theory and the modeling of systems. Englewood Cliffs, New Jersey: Pretence-Hall.
Raff, R.A. and Kaufman, N.C. (1983). Embryos, genes and evolution. The developmental-genetic basis of evolutionary change. New York: Macmillan.
Saussure, F. de. (1959). Course in general linguistics. New York: Philosophical library.
Sebeok, T.A. (1972). Perspectives in zoosemiotics. Hague: Mouton.
Sebeok, T.A. (1976). Contribution to the doctrine of signs. Bloomington: Indiana Univ. RCLSS Publ.
Shannon, C.E. and Weaver, W. (1949). The mathematical theory of communication. Urbana: Univ. of Illinois Press.
Sharov, A.A. (1979). The selection of promising evolutionary directions. Dokladi MOIP (Reports of Moscow Soc. of Researches of Nature, General biology), 1st half-year 1977, 120-122. Moscow: Moscow Univ. Press (in Russian).
Sharov, A.A. (1984). On group selection and its forms. In System principles and ethological approaches in population studies, 36-48 Pushchino, USSR (in Russian).
Sharov, A.A. (1988). Biological metainformation. In Problems of Modern Biology, 1, 15-19. Moscow: deposited in the All-Union Institute on Scientific and Technol. Information, N6710-B88 (in Russian).
Sharov, A.A. (1990). Winter school on biosemiotics. Zhurnal Obshchej Biologii (Journal of General Biology) 51(2), 283-265 (in Russian).
Sharov, A.A. and Kull, K. (1990). Definition and elementary features of self-reproducing systems. Zhurnal Obshchej Biologii (Journal of General Biology) 51(5), 723-730 (in Russian).
Schreider, J.A. (1974). Information and meta-information. Nauchno- technicheskaya informatsia (Scientific-technical information), series 2 (4), 3-10 (in Russian).
Schreider, J.A. and Sharov, A.A. (1982). Systems and models. Moscow: Radio i Sviaz (in Russian).
Stepanov, J.S. (1971). Semiotics. Moscow: Nauka (in Russian).
Teilhard de Chardin, P. (1965). The phenomenon of man. New York: Harper and Row.
Trivers, R.L. (1974). Parent-offspring conflict. Amer. Zoologist 14, 249-264.
Uexkll, J. (1909). Umwelt und Innenwelt der Tiere. Berlin: Springer.
Waddington, C.H. (1968). The basic ideas of biology. In Towards a Theoretical Biology. I. Prolegomena, 1-41. Edinburgh: Edinburgh Univ. Press.
Wilson, E.O. (1980). Sociobiology. Cambridge: Belknap Press.
Zemek, K., Mlikovsky, J. and Socha, R. (1985). Multilevel system of heredity and its ontogenetic and phylogenetic consequences. In Evolution and Morphogenesis, J.A.Novak (ed.), 75-87. Praha.
Zunde, P. (1982). A semiotic approach to information value. In Semiotics 1980 (Proc. 5-th Annu. Meeting of the Semiotic Soc. of America), 587-594. New York: Plenum Press.
Alexei Sharov 09/06/95