Other papers by Alexei Sharov
Semiosis in Self-Producing Systems
Alexei Sharov Department of Entomology, Virginia Polytechnic Institute and State University, Blacksburg, Virginia 24061, sharov@vt.eduAbstract. Cybernetic methodology has reached its limits in the study of life because it ignores the meaning of biological information. Thus it should be augmented by semiotics that studies the meaning and value of signs. According to the pragmatic definition, a sign is a biological adaptation, i.e. a persistent useful function. Usefulness of an action can be measured by its contribution to the reproductive value of an organism in a particular quasi-species. Reproductive values are equal to the components of the left eigenvector of the linearized model of system dynamics. Every organism is a sign, and its life cycle is a continuous process of self-interpretation. Organisms use receptors to predict changing environments. Natural selection is functionally equivalent to perception at the level of lineages. Selective survival and reproduction is analogous to selective excitation of photoreceptors in the eye. Lineages learn how to avoid harmful variation by using developmental constraints, proofreading, dominance, and other mechanisms. If intelligence is defined as the ability to learn, then lineages are intelligent systems, which we did not recognize simply because they are too slow.
Keywords: self-reproduction, value, semiosis, selection, learning.
INTRODUCTION
The idea of informational nature of life became widely accepted after the discovery of the genetic code. Cybernetics and information theory became major tools for solving the mystery of life (1, 2). However, after several decades of work in this direction, the limitations of cybernetics became clear. Cybernetics explains life through the notion of control. But control is only a tool and can be weakened in some situations in order to perform some other functions that appear more important. Thus, a living system is something more than just a control system. The theory of information also failed to explain life because it is most suited for the analysis of context-free languages where any combination of symbols is meaningful. But natural languages (e.g., genetic code or human language) are context-dependent.
The triumph of the Newtonian physics was linked to the idea that we can predict natural events without considering purposes and meanings. But it appears that meanings may be important, especially if we deal with living systems. The first attempt to understand the nature of meaning was done by Charles Peirce (24), who introduced the notion 'semiotics', the theory of signs. He viewed the meaning (sign) as an association between a sign-vehicle and an object via interpretant, which is a change in the interpreter caused by the sign-vehicle that makes him aware of the object. Uexküll (35) developed his theory of meaning independently from Peirce. In this theory he suggested that each living organism develops its own subjective interpretation of its environment, called Umwelt. Each component of Umwelt has specific meaning for an organism (e.g., food, enemy, shelter, etc.) and is associated with one or several functions that support survival and reproduction of the organism. Uexküll was the first to notice the circular nature of biological functions (functional circles). An organism both perceives and modifies external objects as well as parts of its body.
Pattee (23) suggested that meaning emerge in the evolution of self-referenced (semantically closed) systems. He wrote: 'Self-reference that has open-ended evolutionary potential is an autonomous closure between the dynamics (physical laws) of the material aspects and the constraints (syntactic rules) of the symbolic aspects of a physical organization.' According to Pattee, semantic closure requires complementary models of the material and symbolic aspects of the organism. A symbol is a material structure, whose function can not be derived from physical laws, but which is selected for its contribution to the survival of an individual.
Similar ideas were developed by Maturana and Varela (20), who viewed autopoiesis as the crucial characteristic of life and mind. Rosen (27, 28) also emphasized the closed nature of biological organization. He thought that organisms are qualitatively different from machines, because in machines, each function is regulated by another function (which leads to infinite regress of functions). But in organisms all functions are reciprocally closed. Each missing part or function can be repaired or produced by other parts. Mechanisms can be arranged from parts, but organisms can restore and rearrange parts themselves. Thus, organisms can not be built from parts.
Kampis (17) considered life as a semi-closure; the closure is imperfect because of internal freedom in organisms. He emphasized the creative nature of life, which can not be described by deterministic models. Living organisms can invent new functions, whereas deterministic models have all possibilities built in; thus, they can not show any emergent properties. Kampis (17) used models of 'component systems' that are made of numerous small particles of various kinds. But Kauffman (18) thought that emergent behavior can be observed even in deterministic models if the number of possibilities is very large. He modeled self-organization using boolean networks and cellular automata.
There is a consensus on the importance of the symbolic (semiotic) nature of life and its self-referential organization (closure). But there is no agreement on the meaning of terms 'symbol', 'sign', 'self-reference', 'self-reproduction', 'autonomy', 'closure', 'action', 'freedom', and others. Cairns-Smith (3) considered any self-sustaining and self-reproducing system (e.g., crystallization, autocatalysis) a sign or 'gene'. Signs can be isolated or aggregated. Syntax is not present in isolated signs, but may appear in systems of multiple signs. But Rocha (25) thinks that syntax is the essential feature of signs, which is needed for an open-ended evolution. Following von Neumann (37), he views evolution in a traditional Darwinian way as random syntactic changes followed by natural selection. This mechanism of evolution requires the presence of a code that represents an organism. Joslyn (15) hypothesized that all living organisms can be characterized by semiotic autonomy, which means that organisms maintain cyclic relations of perception, interpretation, decision, and action for the sake of increased survival and self-reproduction. Organisms may have various levels of autonomy, and intermediate levels of autonomy are usually more beneficial compared with extremely low or extremely high autonomy. Other authors insist that self-reproduction requires high autonomy. For example, Rose (26) argued that viruses can not self-reproduce because they require a living host cell for interpreting their genetic messages. In contrast, Dawkins (6) thought that strong autonomy is not needed for self-reproduction. Thus, he viewed individual genes and even memes as self-reproducing systems.
Semiotic terminology has a strong anthropomorphic context. Thus, to apply semiotic notions to other living organisms we need to remove all human-specific meanings. This is not an easy task because terms like 'perception', 'action', and 'interpretation' refer mostly to our internal cognitive experience. Also, there is a danger to expand the meaning of semiotic notions too far. For example, Deely (7) thought that even non-living systems are able to interpret signs. He viewed a stone formation that takes on the shape of a dinosaur bone as an interpreter of the bone. But then any physical interaction can be considered an interpretation, and semiotic terminology appears redundant.
In this paper I will review recent publications that indicate the importance of value in semiosis. I believe that the notion of value helps to distinguish semiosis from other processes. Also, it integrates semiotics with evolutionary theory because value represents biological adaptations. The synthesis of semiotics with the evolutionary theory is known as biosemiotics (13, 30).
VALUES AND SELF-PRODUCTION
Economists measure the present value of an investment as a sum of inflation-corrected returns expected in the future. Fisher (11) noticed that the same method could be used to measure the reproductive value of organisms in a population. The reproductive value of an organism of a particular age is equal to its contribution to the growth of the entire population. The rate of population increase is analogous to the inflation rate in economy. For example, eggs have a lower reproductive value than reproducing adults because an adult can produce numerous eggs within a short time, whereas it takes a long time for an egg to develop into adult, and not all eggs will survive to the adult stage.
In a linear model of population growth, the reproductive value of organisms is equal to the left eigenvector of the matrix that describes population dynamics. For example, in the model of Leslie (19), the state of a population is characterized by the vector of age distribution, xt, that shows the number of organisms in each age group at time t. The population dynamics is described by equation
xt +1 = A xt , (1)
where A is the Leslie matrix (Fig. 1)., then the vector of reproductive values, v, is the left eigenvector of matrix A. Newborn organisms have a reproductive value =1 (Fig. 1). As they mature, their reproductive value increases to 2.97, and then decreases to 0.77 in the oldest age group. Vector v satisfies the equation
ATv = lv , (2)
where AT is the transposed matrix A, and l is the largest real eigenvalue, which is the rate of population increase (l = 1.293 for the matrix in Fig. 1). Computation of reproductive values in non-linear systems is more complicated because these systems may have limit cycles or chaotic dynamics (9).
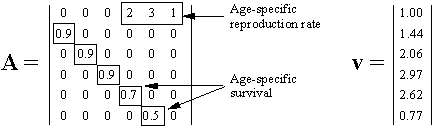
FIGURE 1. A Leslie matrix A and the vector of reproductive values v.
Self-production can be defined as any process by which systems increase their value. For example, development of an organism from egg to adult is self-production because the reproductive value of adults is higher than that of eggs. After laying an egg, the reproductive value of adult female decreases, but the sum of values of an egg and the female is greater than the value of the female before oviposition. An egg is a part of female's self, and egg production is a particular case of self-production. Processes that decrease the value of organisms are usually considered 'external', 'unintentional', or 'random'. They include mortality from various factors (e.g., natural enemies, weather fluctuations), deleterious mutations, etc.
Values exist relative to a particular quasi-species (10), which consists of one or several components with stable relative frequencies. Smith (34) calls them 'virtual species'. Each quasi-species has a specific real eigenvalue l (eq. 2). If a quasi-species changes in evolution, its reproductive values change. For example, if an organism develops effective protection against natural enemies, then the relative value of an egg increases because it gets greater chances to survive to the adult stage. As the value of an egg increases, it becomes beneficial for the parent to invest additional resources into a single egg.
Although quasi-species are discrete, they are not isolated from other systems. I disagree with Rose (26), who does not consider viruses, genes, and memes self-reproducing systems because they require external interpretation. Rose did not notice that the meaning of a viral genome is different for the virus and for the host cell (they are different quasi-species). Thus, the virus does not use a cell as an external interpreter. Instead, it interprets itself by using cell resources. Actually, the cell misinterprets the virus because it considers it a part of its own genome.
A common mistake of evolutionary theory is in considering fitness (l) an objective (i.e., observer-independent) measure of adaptation. Eigenvalues can be estimated for any given model of system dynamics, making an illusion of objectiveness. But subjectivity is hidden in the initial step of model construction. It may happen that in some situations, animals behave differently than it is assumed in the model. If we modify the model to fit their behavior, then all eigenvalues will change (often considerably). Death may seem inevitable in some situations according to our current knowledge. But in a broader sense, death is optional; it results from the inability of an organism to solve its living problems. Thus, fitness shows how smart an animal is in solving its problems. It is as subjective as an IQ measure. Generally speaking, all human knowledge is subjective because it is expressed in a language that represents the history of human experience. We are confident with our language not because it is true but because it was useful in the past.
We can measure the fitness and reproductive values using our model of animal behavior and physiology. But an animal has its own model of its behavior. Of course, animals are not smart enough to solve eq. 2, instead they have a system of values that is based on instinct or learning. In other words, animals can distinguish good from bad, and even have a scale of goodness. It appears that if this scale of values is consistent with system dynamics (i.e., the value changes proportionally with time), then the animal's scale of good/bad is equal to reproductive values. But animals are not as good at distinguishing situations as humans do. They often pool good and bad situations into one category and evaluate them together (e.g., birds do not distinguish wasp-looking flies from wasps).
Humans also rarely assess values using eq. 2. Mostly we use our instincts, intuition, and previous experience in assessing values. But each system of values (if it is consistent with dynamics) corresponds to a specific quasi-species that characterizes life goals. People may see the meaning of their existence in biological survival and reproduction, capital growth, dissemination of their ideas, etc. Human values correspond to quasi-species that people select.
Self-reproduction is a particular case of self-production. Sharov (29) defined self-reproducing systems (SRS) using the formalism of Petri nets. A Petri net has two kinds of components: places and transitions. A place is interpreted as a class of identical objects or as a state of an object. Transitions are processes that either change the state of objects or describe interactions. Arrows connect places and transitions (Fig. 2). Each place may have several tokens on it. A token is interpreted as an object at a particular state. If an object changes its state, then the token is moved from one place to another. If a transition is executed, then the tokens are taken from those places from which arrow lead to this transition, and then tokens are added to the places where arrows go from the transition. The total number of tokens may increase or decrease after the transition is executed. Sharov (29) defined a potential SRS as a subset of places for which there is a sequence of internal transitions (a transition is internal if at least one arrow goes from and to the subset of objects) that increase the number of tokens in all places of this subset. For example, the set of places {x, y} in Fig. 2 is a potential SRS because the sequence of transitions 1, 2, 1, 1, and 2 increases the number of tokens in places x and y. Sharov (29) showed that any potential SRS may become an actual SRS in some favorable environment (e.g., with sufficient resources). He also proved that each potential SRS includes at least one cycle of component-producing processes, a feature that was also mentioned by Csanyi (5).
Self-production processes can be found in non-living systems. For example, a flame of a candle is a self-producing system because it melts the wax. Without melted wax, the flame will go out; thus, the state with more melted wax has a higher value
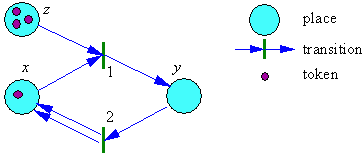
FIGURE 2. A Petri net that represents a simple self-reproducing system {x, y}; z is resource.
for the flame than the state with less melted wax. Self-production of a candle flame normally does not result in reproduction. However, in some special conditions like forest fire flames can reproduce. Crystallization is another example of a self-producing process in a non-living system. But flames and crystals can be viewed also as most primitive living systems, depending on how we define life.
SEMIOSIS GENERATES VALUES
Peirce (24) defined a sign according to its function (correspondence between a sign vehicle, object, and interpretant). But he also emphasized the pragmatic role of signs, i.e., they are useful tools that enhance the performance of living systems. He viewed usefulness as a criterion of truth and meaning of existence. These ideas were further developed by James (14) and Dewey (8), and became a basis of the philosophy of pragmatism. Uexküll (35) added a biological interpretation of signs as components of functional circles that improve the survival and reproduction of living systems at various scales from cells to ecosystems. According to Uexküll, signs are not just a specific category of useful things, but everything that is useful is a sign; in particular, all resources are signs. But the term 'usefulness' remained anthropomorphic, it mostly referred to human intuition on what is useful.
Sharov (30, 33) suggested that usefulness of a process (activity) can be measured by the increment in reproductive value that results from this process. In other words, usefulness of a process is measured by its contribution to the system's survival and reproduction. A sign vehicle can be defined as initial conditions for useful processes. The body of a self-reproducing system (organism) is a sign vehicle that is interpreted in the process of survival and reproduction. The Peircean triadic scheme is still applicabe here: the body is a sign vehicle, inherited characteristics of the body are the interpretant, and the body that is produced (or repaired) is the object (32).
This definition of usefulness changes our understanding of a user. If a carpenter uses a hammer, then the carpenter is a user and the hammer is a tool. But the carpenter uses his arm to hold the hammer. Then the arm is a tool, too. At a closer look, a user is a collection of useful parts connected by useful relations (Fig. 3). Even the brain is a tool. Some people may insist that there is an immaterial soul that uses all body parts. But this assumption is not needed if we accept the idea that an organism is a 'swarming body' (13). Von Foerster (36) noticed that memory is not confined in any particular element of the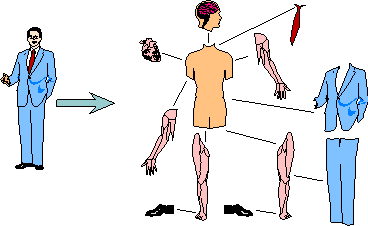
FIGURE 3. A user is a collection of useful parts connected via useful relations.
brain or body; instead, it exists in the form of attractors (eigenstates). Human consciousness has a collective nature which was characterized by metaphors like 'the society of mind' (21) and 'swarm intelligence' (13). Obviously, the value of a whole organism exceeds the sum of values of its parts because parts are linked via useful relations that contribute to the value of the organism.
Interpretation of signs always includes some degree of arbitrariness because it is constrained rather than determined by physical laws (16). At the highest hierarchical level, arbitrariness is represented by the choice of a quasi-species. At this level, the action and the agent are not separated, i.e., in order to select another action, the agent has to change itself by switching to another quasi-species. A self-producing system transfers information about the quasi-species from one generation to another. Inheritance means that the initial state of an offspring belongs to the region of attraction of its parental quasi-species, and a leap to another quasi-species is a mutation. Semiotics can be reformulated by substituting interpreters with quasi-species.
The presence of multiple quasi-species is sufficient for a very primitive form of adaptive evolution. A genealogical line may go through a sequence of quasi-species, each species being a foothold to reach other species. Sharov (31) showed that the Conway's game of life (12) with permutations and a separation rule simulates the emergence and evolution of self-reproducing systems (Fig. 4-5). The separation rule means that if the minimum distance between two configurations is >D, then they become entirely separated as if they have been moved far away from each other (Fig. 4). If D = 4.5, then the spontaneous emergence of the first quasi-species is followed by a mutation to another species with a higher rate of self-reproduction. This model shows that inheritance and adaptive evolution are possible even without a specialized coding mechanism (genome). Coding may develop later as a backup of the functional portion of the system. The 'metabolism first' principle in the origin of life is supported by Cairns-Smith (3), Morowitz (22), and Kauffman (18). The code makes life programmable, which increases the efficiency of further evolution (15).
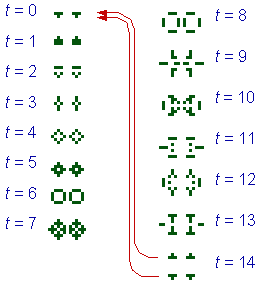
FIGURE 4. A self-reproducing system in the game of life with a separation rule, D = 4.5. At t = 14, two offspring systems become separated (from 31, Ó 1998 IEEE).
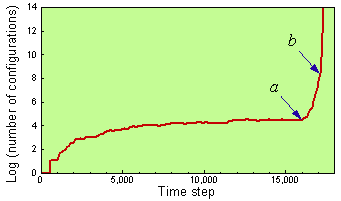
FIGURE 5. Growth of a population of configurations in the game of life with a separation rule (D = 4.5) and with random noise. Arrows show the time when the first (a) and the second (b) self-reproducing species appear (from 31, Ó 1998 IEEE).
NATURAL SELECTION AS PERCEPTION IN LINEAGESThe theory of natural selection is often viewed as a proof that adaptive evolution is a mechanistic process that does not require any subjective or creative factors (6). It is assumed that nature is a mechanical sieve that selects surviving organisms. This interpretation is misleading because selection implies intentionality, but nature has no self-interest in helping or hindering organisms to survive and to reproduce. Hoffmeyer (13) suggested an alternative interpretation of selection as an intentional process at the level of lineages. A lineage probes the environment and tests new adaptations by generating a variety of organisms and copying those organisms that survived. In this interpretation, selection is analogous to perception. Similar ideas were suggested earlier by Kauffman (18, p. 154) who wrote: 'We can think of the population as sending out "feelers" by generating, at random, various mutations. If the mutation occupies a position higher on the terrain, it is fitter, and the population is pulled to the new position.'. Selective survival and reproduction of organisms plays the same role for a lineage as vision (i.e., selective excitation of photoreceptors) for an organism (32).
Perception requires information transfer. A photoreceptor sends an electrical impulse to the brain. It is important that the brain recognizes impulses from individual receptors. If nerve fibers become entangled, then the brain can not associate an incoming impulse with a specific receptor, and the information is lost. Heredity is the information transfer at the level of lineages by which surviving organisms transfer their genetic information to the next generation. If heredity were not accurate enough, then the information would be lost as in the case of entangled nerve fibers.
A possible argument against the analogy between perception and selection is that organisms are able to perceive negative information whereas lineages are not able. The absence of a signal can be a signal for an organism. Darwinism assumes that lineages have no negation operator because organisms that die do not pass any information to surviving organisms. This idea is embedded in the metaphor of 'blind variation'. No matter how many organisms died because of some lethal mutation, the lineage will continue producing this mutation in the future; it does not learn from errors.
Sharov (33) suggested that lineages can use negative information. Eventually, they learn how to avoid errors by creating proofreading mechanisms, dominance, developmental constraints, and other epigenetic tools. The reconstruction of negative information is different in visual perception and selective survival/reproduction. Visual reconstruction is mostly parallel, i.e., it is based on a generalization of signals arriving simultaneously from multiple receptors. But lineages generalize mostly sequential information (but sexual reproduction adds some parallel signals). The persistence of a lineage means that it was successful in avoiding errors in past generations. Its genetical and developmental constraints represent the rules by which errors can be avoided.
Thus, lineages have something like intelligence, but because it is too slow we do not recognize it. Thousands of generations pass before a lineage learns how to avoid a single developmental error. Sexual reproduction can accelerate this process because an organism obtains access to the genetically recorded experience of more individuals. Because selection is perception at a larger scale, there is no qualitative difference between evolution and learning. Both are semiotic processes that increase the relative rate of self-production (fitness) of systems. Fitness, w, is equal to the relative rate of increase of the value, v, of all products of the system
![]()
Then, intelligence (or learning rate) can be measured by the rate of fitness increase:
![]()
According to this definition, intelligence is not fitness (i.e., the ability to solve a given problem). Instead, intelligence is the ability to increase fitness by discovering new solutions of old as well as new problems.
Selective survival and reproduction is the only conceivable way by which the adaptive evolution could start. However, on later stages of evolution, actual reproduction can be replaced partially by virtual reproduction, by which I mean using models and simulations for testing the performance of a system instead of putting the real system at risk. Campbell (4) called it 'vicarious selection' because it replaces natural selection. Vicarious selection is effective only on condition that the modeling strategy is good. If the model fails, then the system dies or suffers big losses in value. In this case natural selection will work until the model is improved. Thus, natural selection is never completely replaced by vicarious selection, but the proportional role of natural selection progressively declines in evolution. Vicarious selection is present in all functions of an organism that involve selective interactions between parts or with the environment: catalysis, selective activation of receptors, transcription and translation of genetic information, apoptosis, immune reaction, interaction of neurons, and others. As natural selection is replaced by vicarious selection, arbitrariness in interpretation of signs moves from the upper hierarchical level to lower levels. As a result, a system may change its actions without any apparent change in its higher-level structure. Thus, it becomes possible to separate the genotype from the phenotype and mind from the body.
CONCLUSIONS
There is a deep similarity between evolutionary, cognitive, and economic processes, which allow us to talk about semiosis and value in any self-producing system. The value of any activity can be measured by its contribution to the rate of self-reproduction in the population. An organism is a sign, i.e., a message to future generations that contains recipes for survival and reproduction. But a lineage (population) is more than a communication channel from one generation to another. It can get new information about changing environments from selective survival/reproduction of its members. Thus, natural selection is equivalent to perception at the level of lineages. Evolutionary systems can be considered intelligent; they use logic and models to reach the goal of stability and self-propagation. However, human intelligence is superior because it is much faster than biological evolution. Intelligence is not an ability to solve a given set of problems, but rather an ability to generate new solutions as well as new problems. The increase of fitness can be used as a simple indicator of intelligence.
REFERENCES
- Ashby, W. R. (1956). Introduction to Cybernetics. Chapman & Hall.
- Bertalanffy, L. von (1969). General System Theory: Foundations, Development, Applications. G. Braziller.
- Cairns-Smith, A. G. (1982). Genetic Takeover and the Mineral Origins of Life. Cambridge University Press.
- Campbell, D. T. (1988). Methodology and Epistemology for Social Science: Selected Papers. Edited by E. S. Overman, University of Chicago Press.
- Csanyi, V. (1998). Evolution: model or metaphor? Pp. 1-12 in: Van de Vijver, G., Salthe, S., and Delpos, M. (eds.), Evolutionary Systems. Kluwer Acad. Publishers.
- Dawkins, R. (1986). The Blind Watchmaker. Longman Scientific & Technical.
- Deely, J. N. (1992). Semiotics and biosemiotics: are sign-science and life-science coextensive? Pp. 47-75 in T. A. Sebeok and J. Umiker-Sebeok (eds.), Biosemiotics: The Semiotic Web 1991, Mouton de Gruyter.
- Dewey, J. (1998). The development of American pragmatism. In: L. A. Hickman and T.M. Alexander. (eds.). The essential Dewey. Vol. 1. Indiana Univ. Press.
- Dubois, D. M. (1998). Emergence of chaos in evolving Volterra ecosystems. Pp. 179-214 in: Van de Vijver, G., Salthe, S., and Delpos, M. (eds.), Evolutionary Systems. Kluwer Acad. Publishers.
- Eigen, M., and P. Schuster. (1979). The Hypercycle. A Principle of Natural Self-Organization. Springer.
- Fisher, R. A. (1930). The Genetical Theory of Natural Selection. Clarendon Press.
- Gardner, M. (1983). Wheels, Life, and Other Mathematical Amusements. W.H. Freeman.
- Hoffmeyer, J. (1996). Signs of Meaning in the Universe. Indiana University Press.
- James, W. (1954). Essays in Pragmatism. Hafner Pub. Co.
- Joslyn, C. (1998). Are life and meaning coextensive? Pp. 413-422 in: Van de Vijver, G., Salthe, S., and Delpos, M. (eds.), Evolutionary Systems. Kluwer Acad. Publishers.
- Juarrero, A. 1998. Causality as constraint. Pp. 233-242. Ibid.
- Kampis, G. (1991). Self-modifying Systems in Biology and Cognitive Science: A New Framework for Dynamics, Information, and Complexity. Pergamon Press.
- Kauffman, S.A. (1993). The Origins of Order: Self-organization and Selection in Evolution. Oxford Univ. Press.
- Leslie, P. H. (1945). On the use of matrices in certain population mathematics. Biometrika, 33: 183-212.
- Maturana, H. R., and F. G. Varela (1980). Autopoiesis and Cognition: The Realization of the Living. D. Reidel Pub. Co.
- Minsky, M. L. (1986). The Society of Mind. Simon and Schuster.
- Morowitz, H. J. (1992). Beginnings of Cellular Life: Metabolism Recapitulates Biogenesis. Yale University Press.
- Pattee, H. H. (1995). Evolving self-reference: matter, symbols, and semantic closure. Communication and Cognition - Artificial Intelligence, 12: 9-28.
- Peirce, C. S. (1955). Philosophical Writings of Peirce. Edited by J. Buchler. New York: Dover Publications.
- Rocha L.M. (1998). Selected self-organization and the semiotics of evolutionary systems. pp. 341-358. in: Van de Vijver, G., Salthe, S., and Delpos, M. (eds.), Evolutionary Systems. Kluwer Acad. Publishers.
- Rose, S. (1998). Lifelines: Biology Beyond Determinism. Oxford Univ. Press.
- Rosen, R. (1985).Anticipatory Systems: Philosophical, Mathematical and Methodological Foundations. Pergamon Press.
- Rosen, R. (1991).Life Itself: A Comprehensive Inquiry into the Nature, Origin, and Fabrication of Life. Columbia University Press.
- Sharov, A. A. (1991). Self-reproducing systems: structure, niche relations and evolution. BioSystems, vol. 25, pp. 237-249. Get a reprint! (PDF), Short version (HTML)
- Sharov, A. A. (1992). Biosemiotics: a functional-evolutionary approach to the analysis of the sense of information. Pp. 345-373 in: T. A. Sebeok and J. Umiker-Sebeok (eds.), Biosemiotics: The Semiotic Web 1991. Mouton de Gruyter. Read it! (HTML)
- Sharov, A. A. (1998). Signs and Values. Pp. 677-682. Proceedings of the 1998 IEEE ISIC/CIRA/ISAS Joint Conference. Gaithersburg, MD. September 14-17. IEEE.
- Sharov, A. A. (1998). From cybernetics to semiotics in biology. Semiotica 120 (3/4), 403-419. Read a short version (HTML)
- Sharov, A. A. (1999). Umwelt-theory and pragmatism. Semiotica (in press).
- Smith J. H. (1998). Canonical ensembles. Pp. 141-153 in: Van de Vijver, G., Salthe, S., and Delpos, M. (eds.) Evolutionary Systems. Kluwer Acad. Publishers.
- Uexküll, J. von (1982). The Theory of Meaning. Semiotica 42(1), 25-82.
- von Foerster, H. (1969). What is memory that it may have hindsight and foresight as well? Pp. 19-65 in S. Bogoch (ed.), The Future of the Brain Sciences. New York: Plenum Press.
- von Neumann, J. (1966). Theory of Self-Reproducing Automata. Edited and completed by A. W. Burks. University of Illinois Press.
Alexei Sharov 11/22/99