Signs and values
Virginia Polytechnic Institute and State University, Blacksburg, VA 24061-0319
Abstract
I consider the value (usefulness) as the most important characteristic of a sign. Values exist only in self-reproducing systems which satisfy the criterion of semantic closure. They can be estimated in a linear model of a system as left eigenvector. The value of any relationship among system components is equal to the contribution of this relationship to the overall value of the system. Autocatalytic systems are most primitive signs that are in a permanent process of self-interpretation. Signs are produced only if organisms expect to get a return from this production (increased self-reproduction rate). In the same way, organisms interpret signs only if they expect to increase their value from interpretation. Organisms are investors that distribute their resources into various communication channels and their profit is self-reproduction. Human values depend more on the propagation of life styles (memes) rather than biological reproduction, and selection goes mostly at a sub-organism (mental) levels. In evolution, signs develop a hierarchical structure via metasystem transitions. Two mechanisms of metasystem transitions are possible: multiplication with subsequent cooperation, and symbiosis. Living cells apparently originated from a symbiosis of self-reproducing linear polymers with self-reproducing membrane spheres. Hierarchical systems are integrated via communication of their components. Cooperation is evolutionary stable only in encapsulated systems in which communication is restricted. Value conflicts are resolved in evolution by limitations of neutral variation of components.
1. Introduction
What is life? This question first explicitly formulated by Schrödinger (1944) still has no clear answer. The problem is no longer to get a formal distinction between living and non-living objects which is as meaningless as attempts to find a monkey that gave birth to a first human. More interesting is to examine formal models of living systems that may indicate relations among various characteristics of life. For example, there is a question to find sufficient conditions for an open-ended evolution. Or it is interesting to examine general properties of self-reproducing systems (Von Neuman 1966, Sharov 1991). This area of study is known as artificial life (Langton 1989).
Communication is one of the most important characteristics of life (Hoffmeyer 1997). Examples of communication in living systems are: transfer of parent genome to offspring organisms, chemical and acoustic signals, etc. Mechanisms of interpretation vary from simple transcription and translation of DNA to complex chemo-electrical processes in the brain. Biosemiotics is a new interdisciplinary branch of science that studies communication and interpretation of signs in living organisms (Sebeok 1972, Sharov 1992, Hoffmeyer 1997).
In this paper I attempt to link communication in living systems with their ability of self-reproduction. I show that self-reproduction always require communication, and any communication is a part of a larger self-reproduction cycle that represents semantic closure (Pattee 1995). This brings together biosemiotics with the theory of adaptation, and indicates that the most important characteristic of a sign is its value (usefulness).
2. Self-reproduction
Intuitively, self-reproduction means that a system is able to produce multiple copies of itself some time later. Sharov (1991) formalized this idea using the Petri nets. A Petri net consists of places and transitions between places. A place is interpreted as a state of objects, and transitions may transfer an object from one state to another. Also transitions may merge several objects into one, or split an object into several parts. Sharov (1991) defined a potential self-reproducing system as a finite subset of places for which there is a sequence of transitions that increase the number of objects in all places of this subset.
This definition has not been generalized for continuous phase spaces. Apparently, a self-reproducing system in a continuous space is similar to attractor. But the notion of attractor has to be modified because trajectories are branching.
Self-reproduction always require information transfer because a parent system produces offspring in specific initial conditions that ensure a return to the state of the parent system. In continuous phase spaces, offspring systems should be produced within the same domain of attraction. Accidental transfer into the domain of another attractor is equivalent to mutation.
Von Neuman (1966) noticed that self-reproduction itself is not a sufficient condition for an open-end evolution. Simple autocatalytic systems (e.g., nuclear reaction) usually do not evolve. According to Von Neuman (1966), only those systems can evolve in which self-reproduction is based either on self-examination or coding. However, it was shown that some autocatalytic networks can evolve even without self-examination or coding (Bagley et al 1991).
I found evolutionary processes in simple cellular automata with separation rules (Sharov, unpublished). The system is based on the Conway�s game of life in which the following separation rule is applied at each time step: if the minimum distance between 2 configurations is >D, then they become entirely separated as if they were moved far away from each other. Because of the separation rule, the trajectory is branching. Separation rules solve the "overcrowding" problem that limits self-reproduction in traditional cellular automata. The dynamics of cellular automata with a separation rule resembles the dynamics of autocatalytic networks. For the game of life with a separation rule (D = 4.5) I found several self-reproducing attractors with different intrinsic growth rates. An example of a self-reproducing configuration is given in Fig. 1.
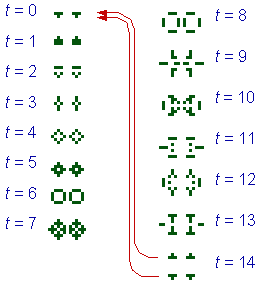
Fig. 1. A self-reproducing configuration in the game of life with a separation rule, D = 4.5.
If random modifications of existing configurations (i.e., noise) are added to the model, then self-reproducing configurations may develop spontaneously and then evolve in the direction of increasing intrinsic growth rates (Fig. 2).
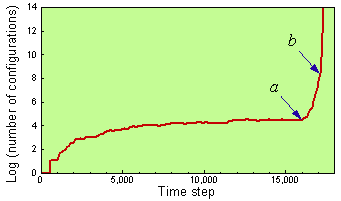
Fig. 2. Growth of the population of configurations in the game of life with a separation rule (D = 4.5) and with a random noise. Arrows show the time when the first (a) and the second (b) self-reproducing species appear.
Evolution is based on transitions between self-reproducing attractors (mutations). Von Neuman (1966) was right that self-examination and coding are mechanisms that simplify transitions between attractors. But these mechanisms require complex organization that can not be expected in systems which existed at the very beginning of life. Apparently, life started from much simpler systems that had no coding. But the complexity of these systems should be sufficient for possible transitions between attractors.
The distinction between coding and no coding is the same as between a word that consist of several characters and a hieroglyphic. Primitive self-reproducing systems transfer information to their progeny as a hieroglyphic (each hieroglyphic corresponds to a self-reproducing attractor). More advanced organisms learned how to split a hieroglyphic into characters, and this invention accelerated further evolution.
The boundary between life and non-life can be drawn at that level of complexity where transitions between self-reproducing attractors become more frequent than spontaneous emergence. Obviously, this boundary is fuzzy and may depend on external conditions because some processes that are impossible on Earth may be possible in other places in the Universe and vice versa.
3. Values
Traditional physics never studied values (i.e., usefulness) of objects. But the notion of value is very important for understanding the phenomenon of life. Components of the environment may have positive or negative value for an organism. For example, resources have positive values, and dangerous objects (e.g., enemies) have negative values. Values can be applied also to various kinds of activity: eating, sleeping, moving, growing, reproducing, etc. By evaluating objects and processes, an organism subjectively interprets the world and itself. Uexküll (1940) used the word "Umwelt" to designate this interpreted world. The word "Umwelt" means environment in German, but Uexküll modified its meaning. Umwelt can be viewed as a model of the world that is used by an organism in its activities. For example, an ant and a cow perceive the same environment (meadow) in a very different way. Grass stems are food for the cow and bridges for the ant.
Usefulness is not a quality but a relation between an object and user. But how to define a user? At a closer look, a user is nothing but a collection of useful objects. Organs are tools that are used by an organism for performing specific functions, but there is nothing in the organism besides organs. There is no homunculus who determines the value of organs. Thus, the user is just a set of relations between useful parts.
Obviously, not all kinds of relations can be considered useful. Some relations may destroy the system. Relations are useful only if they preserve and augment the same relations in the future, i.e., if these relations are self-reproducing. This idea was first formulated by Pattee (1982, 1995) and was called "semantic closure". Pattee (1995) defined semantic closure in terms of matter-symbol relationship. Matter is the part of our experience represented by observer-independent and universal physical laws. In other words, matter is what remains constant. Symbols, are results of measurement, and they represent initial conditions which may vary. Usefulness is always associated with initial conditions that are beneficial for achieving goals and, eventually, for population growth. Thus, the notion of usefulness can not be handled by physics. Universal laws can not be changed, by definition; therefore, the notion of usefulness is not applicable in physics. It is necessary to note that Pattee (1995) considered physics as an abstract theory without any applications; apparently, he considered applications as engineering.
Semantic closure is a new criterion for autonomy (or wholeness) of systems. A set of elements connected by relations is autonomous only if it is semantically closed, i.e., it reproduces itself in the future. The value of each component or relation in an autonomous system corresponds to its contribution to the ability (or probability) of the system to reproduce itself.
The notion of value was introduced to biology by Fisher (1930). He defined reproductive value of an organism as its contribution to the growth of the entire population. For example, eggs have a smaller reproductive value than adults because adults can easily produce multiple eggs, but it takes a long time for an egg to develop into adult. Adults at the beginning of their reproduction period usually have the highest reproductive value; but as they grow older, their reproductive value declines. Fisher (1930) took the notion of value from economy because he noticed the analogy between exponentially growing population and exponentially growing market. To estimate reproductive value for organisms of various age, a linear model of the population with age structure should be used (e.g., a Leslie model). Reproductive value is equal to the left eigenvector of the matrix of this model that corresponds to the highest positive eigenvalue. If the dynamics of the system is non-linear, then the time scale of the model should be increased until the dynamics will become close to linear.
In simple organisms that are not able to learn from their individual experience, natural selection is the mechanism that maximizes the value of organisms at each step in the life cycle. For example, if an adult insect has a choice to lay 1 egg with a reproductive value of 1.0 or to lay 10 eggs with a reproductive value of 0.2, then natural selection will favor the second option because the total value produced is greater. Higher animals can estimate the value of their products before natural selection will take place. Thus, they are able to optimize their behavior faster than organisms without such mental abilities.
Fisher (1930) defined values only for entire organisms, but this definition can be extended to the parts of an organism and to relations between parts. The value of a part (or relation) is equal to its contribution to the process of self-reproduction. For example, the value of the resource is equal to the gain in reproductive value of an organism that captured this resource unit.
4. Signs
Biologists may ask why to use semiotic terminology in simple population models? In particular, why to talk about semantic closure instead of self-reproduction? "Self-reproduction" seems to be a convenient term that does not have uncertainties associated with signs or semantics. But this simplicity is illusive; self-reproduction includes the word "self" which comes from the field of semiotics rather than physics or biology. In the process of self-reproduction, an organism defines itself; in other words, self is what is preserved in the process of self-reproduction. Self-reproduction is simultaneously a process of self-measurement, self-interpretation, and communication from parents to offsprings.
Communication is the essential characteristic of living systems (see Introduction). In this section I discuss the nature of signs which are objects used for communication. Peirce (1955) defined a sign as a triadic relationship between a sign vehicle (representamen), an object, and interpretant which is a representation of the object in human mind invoked by the sign vehicle. The interpretant is a mental model of an object. Bacteria are not able to build mental models of objects but they can build material models of themselves, i.e. their offspring. Genome can be viewed as a sign vehicle that is interpreted in offsprings. It tells offspring organisms how to develop, survive, and reproduce. The message is true because it was verified by natural selection in numerous generations. In simple autocatalytic systems, genome is not represented by a specialized structure (e.g., DNA), and the entire system can be viewed as a message (i.e., sign).
This analysis shows that a sign is an object that is involved in the process of self-reproduction, i.e., an object that has value. Resource is a primitive sign; it is recognized by the system in the process of utilization. Organisms produce various signs: offspring organisms, sounds, chemical signals, etc. Selection favors those organisms which produce signs with a higher total value.
Normal communication requires that signs have positive values both for a producer and receiver (Sharov 1992). An organism spends its resources to produce a sign only if the sign has value, i.e., it increases the rate of self-reproduction either directly or indirectly. In the same way, the receiver never interprets a message unless it expects to increase its fitness after interpretation. Here I mean expectation in a broad sense including evolutionary (unconscious) expectation. Only in higher animals and humans expectation becomes conscious.
But in some cases, the value of signs may be negative. For example, some predators may intercept signals produced by their prey. In this case, the value of a sign is negative for the producer. Other predators may emulate signs that attract their prey. In this case, the value of a sign is negative for the receiver. But negative values are not normal. If a sign has negative value too often, then organisms will simply avoid using it.
Human signs also have values, but this value is no longer connected with biological reproduction. Human evolution is driven more by the propagation of life styles (memes) rather than by propagation of genes. Memes are associated with specific human relations (e.g., ethical, religious, educational, etc.). The value of texts is associated with propagation of these relations. Peirce (1955) described only a half of the life cycle of a sign, i.e., the process of perception and recognition. He did not analyze the process of sign production which closes the cycle (semantic closure). According to Pattee (1995) each sign participates in a larger system with semantic closure.
5. Metasystem TransitionsThe major trend in the evolution of signs is the growth of their hierarchical structure. Simple autocatalytic systems represent a single hieroglyphic without internal structure. More advanced systems develop a hierarchical signs which are similar to signs in a human language. For example, elements of genetic language are nucleotides, triplets, codons, exons, introns, genes, chromosomes.
Development of new hierarchical levels in autonomous systems was called metasystem transition (Turchin 1977). Metasystem transition starts with duplicating of original systems and ends with establishment of a new semantic closure (Turchin used the term "control"). For example, multicellular organisms originated via duplication of cells that stayed together and cooperated.
Symbiosis is another mechanism of metasystem transitions which was not mentioned by Turchin (1977). Several non-similar systems may start cooperating, and this cooperation represents semantic closure at a higher level. For example, lichens are simbiotic organisms that originated from fungi and algae; eucaryotic cells originated from a symbiosis of several types of procaryotic cells.
When two autocatalytic systems cooperate, they produce resources for each other. Resources, as we have seen in the previous section, are primitive signs. Thus, cooperation is a semiotic relationship. Each component has a double interpretation in such a system. First, it is self-reproducing on its own (self-interpretation), and second, it produces signs that are interpreted by another component.
The major obstacle on the way of cooperation is possible evolutionary instability. Let us consider cooperating chemical species A and B that produce resources for each other. Species A may "mutate" into a selfish species A1 which will use resources produced by species B without providing help to the species B. As a result, the cooperation between species becomes broken.
Cooperation is evolutionary stable only if specific restrictions are applied on communication (resource exchange). I call these restrictions "encapsulation". For example, several representatives of species A and B may form small groups, so that communication occurs only among members of a group. If a selfish mutation destroys communication within a group, then this group will become less competitive and eventually will be eliminated in the process of group selection. Thus, encapsulation makes a metasystem transition possible. Hierarchical systems have several levels of encapsulation and this makes them more stable than systems that have no communication restrictions.
The theory of metasystem transitions can be used to outline possible initial steps in the evolution of life. The major components of all living organisms are 1-dimensional and 2-dimensional polymers (linear polymers and membranes). Morowitz (1992) indicated that membranes should appear at the very beginning of life. Both linear polymers (Fig. 3A) and membranes (Fig. 3B) may grow by adding monomers at the ends or inserting them. These polymers may reproduce themselves via breaking into smaller portions after disturbances. The sequence of monomers can not be inherited in this way.
Apparently, the first metasystem transition was a symbiosis of linear polymers with membranes (Fig. 3C). Membranes provide encapsulation which is necessary for evolutionary stability (see above). Linear polymers may benefit from encapsulation because (1) they become protected from external disturbances, and/or (2) the rate of autocatalysis increases due to a higher concentration. Membranes may benefit from linear polymers because they may grow by incorporating molecules produced by linear polymers. Mutual benefits create a semantic closure.
Further steps can be viewed as follows. The number of linear components in a cell increases (Fig. 3D), and inheritance is achieved by transfer of at least one molecule of each kind to the offspring cell. At the next step, linear molecules "learn" how to build a complimentary copy by their side (Fig. 3E). This new mechanism of self-reproduction is faster than attaching monomers at the ends and, most important, it may transfer information about the sequence of monomers. The sequence of monomers may determine polymer folding, and hence, its catalytic activity. Some folded polymers may be able to synthesize other cell components. Thus, information transfer to daughter cells can be limited to a set of irreplaceable elements. These irreplaceable elements represent the "genome". Synthesis of cell components should be regulated; thus, molecules that modify catalytic synthesis are needed (Fig. 3F). If a folded catalyst "learn" how to pull the modifier molecule through itself, then it may modify the sequence of its product according to the sequence of the modifier molecule (Fig. 3G). This explains the origin of the genetic code.
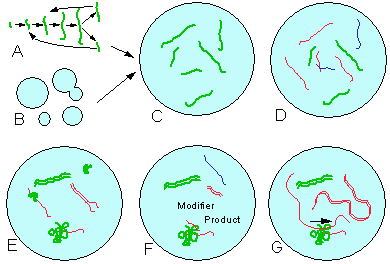
3. Hypothetical scheme of the origin of life. A and B: self-reproducing linear polymers and membranes; C: symbiosis of linear polymers and membranes; D: the number of components increases; E: complimentary duplication, folding, and catalysis; F: regulated catalysis; G: coded catalysis (genetic code).
Each self-reproducing component in a hierarchical system defines its own values. Thus, there is also a hierarchy of values. A message may have value for several components. For example, growth hormones stimulate proliferation of individual cells and thus, they have a positive value for a cell. But the same hormones may change the morphology and function of a multicellular organism. These changes may be beneficial or harmful. Also, growth hormones may cause cancer which is fatal for an organism.
The value of an element at a higher level may be reduced if there is a conflict of values among its components. Thus, natural selection favors those hierarchies in which conflicts are minimized. The fitness landscape of components usually does not have a single sharp peak. Instead, there is a region of neutrality where fitness is almost the same. Thus, the super-system may examine neutrality regions of its sub-systems and find points where conflicts between sub-systems are minimized. The system creates a higher-level value without damaging lower-level values. The ability to create new values on the top of existing values is the basis of a free will.
References
Bagley, R. J., J. D. Farmer, and W. Fontana. 1991. Evolution of a metabolism. Pp. 141-158. In: C. G. Langton, C. Taylor, J. D. Farmer, and S. Rasmussen (eds.), Artificial life II, SFI Studies in the Science of Complexity, vol. X. Addison-Wesley, Redwood City, CA.
Fisher, R. A. 1930. The genetical theory of natural selection. Clarendon Press, Oxford.
Hoffmeyer, J. 1997. Signs of meaning in the Universe. Indiana University Press, Bloomington.
Langton, C. G., ed. 1989. Artificial life. SFI Studies in the Science of Complexity, vol. VI. Addison-Wesley, Redwood City, CA.
Morowitz, H. J. 1992. Beginnings of cellular life: metabolism recapitulates biogenesis. Yale University Press, New Haven.
Pattee, H. H. 1982. Cell psychology: An evolutionary approach to the symbol-matter problem. Cognition and Brain Theory 5: 325-341.
Pattee, H. H. 1995. Evolving self-reference: matter, symbols, and semantic closure. Communication and Cognition - Artificial Intelligence, 12, 9-28.
Peirce, C. S. 1955. Philosophical writings of Peirce. Edited by J. Buchler. Dover Publications, New York.
Schrödinger, E. 1944. What is life? The physical aspect of the living cell. Macmillan, New York.
Sebeok, T. A. (1972). Perspectives in zoosemiotics. Mouton, The Hague.
Sharov, A. A. 1991. Self-reproducing systems: structure, niche relations and evolution. BioSystems, 25: 237-249.
Sharov, A. A. 1992. Biosemiotics: a functional-evolutionary approach to the analysis of the sense of information. Pp. 345-373. In: Sebeok, T. A. and J. Umiker-Sebeok (eds.), Biosemiotics: The Semiotic Web 1991. Mouton de Gruyter, Berlin.
Turchin, V. F. 1977. The phenomenon of science. Columbia University Press, New York.
Uexküll, J. von 1940 (1982). The Theory of Meaning. Semiotica 42, 25-82.
Von Neumann, J., 1966, Theory of self reproducing automata. Univ. of Illinois Press, Urbana.
Alexei Sharov 5/06/98