Other papers by Alexei Sharov
Towards the semiotic paradigm in biology
Virginia Polytechnic Institute and State University, Blacksburg, VA 24061
Contemporary biology considers living organisms as complex machines; and the common belief is that their dynamics can be explained by studying each process and each component separately. If this knowledge is integrated into a model, it can predict the dynamics of organisms. Although biologists continue studying tiny details in animals and plants, it becomes clear that we do not have tools to integrate this knowledge. Integrative models can not be built because: it is impossible to get exhaustive knowledge about all system processes and components, and the model behavior does not converge to the dynamics of the real system as we incorporate new information into the model. It is impossible to get exhaustive knowledge because any living system is unique at any moment. The dynamics of models do not converge to the observed system dynamics because the role of processes may change. Biological processes are usually studied in the order of their decreasing importance. However, the role of processes may change either spontaneously or because of environmental changes. Thus, some unknown processes would become more important than those processes that were already studied. The model could be thoroughly validated, but it still may fail in further studies.
Extreme reductionism was replaced by cybernetics that uses generalized descriptions, i.e., models that do not represent all available knowledge about the system. The concept of information is a very useful aggregate characteristic of biological systems (Wiener 1962). For example, adrenaline can be considered as a signal to which liver cells respond by releasing glucose. It does not matter that the chemical mechanism of cell response is known only partially. We can simply consider the cell as a black box with adrenaline as input and glucose as output and ignore everything that happens inside. Then, the whole system can be modeled as a set of interconnected black boxes with information flows between them. This kind of models can be used to examine system stability and to optimize control in order to reach specific goals.
The major limitation of cybernetics is the assumption that signal-response relationships are pre-determined. The origin of this relationship is not considered. For example, the response of cells to insulin is assumed to be the same and independent from cell environment and history. The first impression is that this problem can be easily fixed by adding input factors and/or by enlarging the number of internal states of the system. This could be a solution if all possible inputs and their interpretations were known. But then no novelty could be expected in the evolution of life, and no free will can be assumed in humans (Kampis 1991). Cybernetic methods may work well in predicting behaviors of biological systems in short time-scale, but they cannot be applied to long-scale biological processes, like evolution, in which probabilities cannot be estimated. For example, it is impossible to calculate the probability of the origin of man from monkeys. Cybernetics does not capture the specifics of life because its models equally work in living and non-living systems. Thus, it is clear that cybernetics failed as a general theory of life.
Biosemiotics is an alternative current of thought which does not accept deterministic or probability models. It denies all forms of objective (i.e., observer-independent) determinism and develops a concept of subjective determinism which is expectation or goal. This is a pivot change in scientific mentality which is not easy to accept in traditional science. Thus, biosemiotics is still an alien in biology.
Biosemiotics have roots both in biology and semiotics which is a theory of signs. Jacob von Uexküll (1940) can be considered the founder of biosemiotics, although he did not use this term. He proposed the notion of ‘Umwelt’ which is the world seen through the eyes of an animal. Each animal associates external objects with some meaning which is specific to its habits. For example, an ant considers plant stems as a path to its food area in the flower, but a cow considers them as food. Sebeok (1972) adopted semiotic methods and terminology to describe signification in animals and called his theory ‘zoosemiotics’. Later signification was described in plants, and the term ‘phytosemiotics’ appeared (Krampen 1981). Sign processes penetrates the entire body of an organism. The DNA molecule codes the sequence of amino acids in proteins, which in turn may be signals for various kinds of actions at a cell or organism level. Cells communicate with each other using signal molecules (hormones, mediators). Sebeok (1976) suggested the term ‘endosemiotics’ for signification among parts of the body. The term ‘biosemiotics’ apparently was first used by Stepanov (1971). Deely (1982) considers biosemiotics as a branch of semiotics that addresses all signification processes going in living organisms including humans.
An alternative approach is to consider signification as the fundamental property of living systems that can be taken as a definition of life (Pattee 1972, 1995, Sharov 1992). Hence, biosemiotics can be viewed as a root of both biology and semiotics rather than a branch of semiotics. Human semiotics can be fully understood only after analyzing the evolution of signification processes in living organisms starting from the origin of life. Thus, biosemiotics is opposed both to physicalism in biology and to anthropomorphism in semiotics.
Organism as a Message. Living organisms have internal self-description written in a DNA form. This description comes from previous generations and summarize the experience of all ancestors in the art of surviving. Thus, an organism has a dual nature: it stands for itself and it is also a message sent from all previous generations to all future generations (Calow 1978, Hoffmeyer 1997). This duality is the essential feature of life which makes biological evolution possible. Differential survival and reproduction of organisms is a semiotic process which incorporates present into the future. Hoffmeyer (1997: 16) characterizes life as survival in a coded form. Messages that provide better recipes for surviving are reproduced together with organisms whereas messages with poor instructions disappear together with their bearers.
Coding is based on conventionality. For example, the correspondence of DNA triplets to amino acids in proteins does not follow from any physical or chemical laws; it is a semiotic correspondence. Peirce characterized this arbitrariness by the term ‘habit’ because habits result from a free choice rather than forced externally. Life is full of habits exhibited at various hierarchical levels from molecules to organisms. Most habits in animals are coded in the DNA. Also, higher animals and humans developed a new social coding that allows to transfer acquired habits.
A message requires an interpreter, and the genetic code is interpreted by each cell in the body. The offspring organism is supplied with both the code and interpreter that can read this code and build an organism from it. The processes of interpretation of the genetic code (organism building) are called epigenetic processes. Thus, heredity always has genetic and epigenetic components.
Semiotic Triads. Peirce suggested a triadic scheme for a sign which is the relationship between a sign vehicle (e.g., the dinosaur bone), an object (the dinosaur), and interpretant (the recognition of the dinosaur from the bone by paleontologist) (Deely 1992). Hoffmeyer (1997) applies triads to genetic code by considering the DNA as a sign vehicle, fertilized egg as interpretant, and ontogenetic trajectory as object. Despite obvious differences between these two examples, they both satisfy the conditions of a sign relationship. In both cases the sign vehicle triggers the process of building another object: the mental model of the dinosaur or the material model of an organism. But if an organism is the model then the model of what? I think that any organism is a model of its ancestors. And as with any other model, it may slightly deviate from the original.
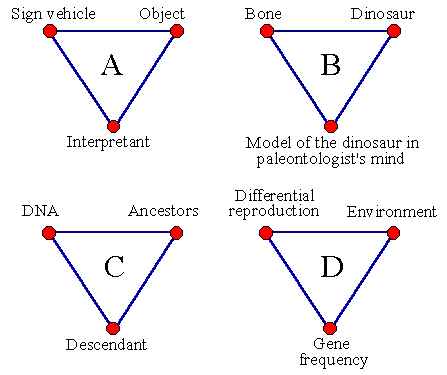
Fig. 1. General sign triad (A) and its application to: B, recognition of the dinosaur by paleontologist; C, construction of a descendant organism that is a model of the ancestor coded in the DNA; and D, change in gene frequencies in a population due to selective mortality.
It seems that the analogy between human signs and genetic signs would be more clear if we modify the triadic scheme presented by Hoffmeyer (1997). The ontogenetic trajectory of the descendant organism is interpretant rather than an object (Fig. 1). Because DNA represents ancestor organisms, these ancestors should be considered as an object. Hoffmeyer (1997) suggested that the fertilized egg is the interpretant which is not correct because the egg is the interpreter. Peirce did not include interpreter into the triad; thus the fertilized egg is not shown in Fig. 1.
Hoffmeyer (1997) applied a triadic scheme to the natural selection which causes changes in gene frequencies. He considers a lineage (ancestor-descendant sequence) as interpretant, and the ecological niche as a sign vehicle that indicates that the DNA pool in the population should be modified (DNA pool is an object). I again disagree with this interpretation. The lineage is the interpreter rather than interpretant, and the ecological niche is an object rather than the sign vehicle. The lineage ‘sees’ the environment through selective reproduction of its organisms in the same way as human eye sees the world through selective activation of photo-receptors. Thus, differential reproduction is a sign vehicle, environment is the object and the change in gene frequencies is the interpretant (Fig. 1D). Gene frequency in the population is a model of the environment which contains recipes for survival in this environment. If the environment changes, then gene frequencies are adjusted by the lineage so that they will again correspond to a better algorithm of survival and reproduction in these new conditions.
Hoffmeyer (1997) offers a new interpretation of the evolutionary process. He points out that the term “natural selection” is confusing and misleading because it is the lineage rather than Nature who selects most successful organisms for reproduction in future generations. According to Darwin, evolution is the result of 3 factors: heredity, variability, and selection. Hoffmeyer’s interpretation integrates all these factors into one because the lineage selects organisms for future reproduction through mechanisms of heredity and variability.
Triadic schemes like those shown in Fig. 1 can be helpful for drawing analogies between human signs and signs used by animals and plants. However, they do not capture at least 2 essential features of signs and thus can be misleading. First, the interpreter is not explicitly present in the scheme and is often confused with the interpretant. Second, there is always a reason for interpreting signs which means that the interpreter expects to gain some value from a sign. If values are ignored then any physical interaction can be considered a sign and the notion of sign will loose its content.
The question of value is very important because it is related to the question whether semiotic relationships can be found in non-living nature. Deely (1992) thought that signs exist both in living and non-living nature. He considered the following example of a sign that does not involve living organisms. When a bone of the dinosaur is fossilized, then the geological stone formation which used to be a bone is the interpretant of the bone, and the interpretation points to the dinosaur as object. I agree that the geological formation has changed in its interaction with the bone, but I see no sign relationships here. Stone formation does not expect to gain any value from interpreting the sign. Moreover, the bone itself does not point to the dinosaur. Only a trained paleontologist can associate the bone with his mental model of dinosaurs which does not originate from this particular bone.
Swarming Body. Sign relationships may exist only if there is ‘somebody’ or ‘self’ who interprets signs. Thus, the notion of ‘self’ need to be clarified. Traditional semiotics had no problem defining the interpreter because it was always a human being. But in biosemiotics, the term ‘interpreter’ is not that clear. Any organism, a bee colony, a cell in the multicellular body, and even a molecule (receptor) can be considered an interpreter. The problem is that interpreters are hierarchically organized and it is important to distinguish semiotic processes going at various levels.
We (humans) consider our consciousness as something united. When we have a choice we select only one option, for example, we cannot simultaneously go to the right and to the left. Hoffmeyer (1997) argues that this unity is illusory. He introduces concepts of ‘swarming body’ and ‘swarming intelligence’ which mean a decentralized self. If the self was centralized then there would be just one hierarchical level of selves which contradicts to numerous evidence of endosemiosis. According to the ‘swarming body’ concept, semiosis is always organized hierarchically. If subunits of an organism were not able to interpret signs, then they could be controlled only mechanically. Thus, the alternative to the ‘swarming body’ is a mechanism.
After a close look, our mind also appears to be a collection of ‘smaller minds’. Minsky (1986) views human mind as a society of small subunits which perform isolated tasks. Some of them are responsible for recording and analyzing external signals, some of them are responsible for memory, and so on. All these mind elements control the same body, and thus, they have to come to an agreement before the action is taken. The agreement can be achieved by something like voting. A similar idea was developed by Gazzaniga (1985).
Umwelt. The notion of Umwelt was introduced by Uexküll (1940) for a subjective world as it is perceived and modeled by an organism. Hoffmeyer (1997) emphasized the activity of organisms in building their Umwelts. Recipes for building Umwelts are included in the genotype and are transferred selectively to subsequent generations. Thus, the evolution is not just a change in the shape of organs but rather a change in semiotic relationship with the world. The major evolutionary trend is the selection in favor of more complex Umwelts that increase the semiotic freedom of organisms.
According to Hoffmeyer (1997), consciousness appear when an organism discovers that it is the builder of its own Umwelt. Because of this circular reference, humans obtained freedom to reinvent semiotic relationships with the world, which enormously speeds up the evolution of Umwelts. But freedom creates ethical problems which we will discuss below.
Value. One of the basic problems in biosemiotics is understanding how ‘something’ becomes ‘someone’ Hoffmeyer (1997). In other words, what is the origin of subjectivity, or how a system can start interpreting the world? Action is the only possible predecessor of interpretation. In primitive organisms, interpretation is always reduced to some simple action. For example, the sound of a bat means ‘fall down’ for a moth, and a source of light means ‘move here’ for unicellular algae. Organisms also exhibit numerous spontaneous actions which are not induced by any external signal. The question is: what makes organisms perform all these actions? According to physics and chemistry, organism activity is predetermined in its structure. But this is only a half of an answer or, better to say, not an answer at all because the next question is why organisms have this specific structure. Action and structure are two sides of the same thing which can be called ‘organization’.
The explanation of the origin of organization comes from the theory of evolution which in its most general form states that action and structure may form a self-stimulating loop based on self-reproduction (Conrad 1983). As a result, action and structure are integrated into an organism.
The value of system A relative to system B is a measure of the ability of system A to produce systems B. Value depends on how many systems B are produced and on how soon they are produced. It can be measured by the number of systems B produced at various times weighted by a discount function which decreases exponentially with time. This definition of value is analogous to the notion of present value in economics which is the inflation-adjusted total net benefits obtained from taking a specific action. It is also equivalent to the notion of reproductive value in population ecology (Pielou 1974).
An organism can be viewed as an investor whose profit is self-production (Sharov 1992). To maximize the rate of self-production, it should produce systems with maximal total value relative to itself. For example, an adult insect may immediately spend its entire energy on laying eggs and then die, or it may lay a few eggs and continue living. If the decrease in the value of this insect after laying an egg is smaller than the value of an egg, then it is beneficial to continue laying eggs. However, if the decrease in the value of adult insect is greater than the value of an egg, then it is beneficial to cease laying eggs so that the insect would be able to produce more eggs in the future. Thus, every process in living systems is optimized within physical and biological constraints (Rosen 1967).
Altruistic behavior in social insects (e.g., bees and ants) is investment into relatives. The genotypes of related organisms are similar. Hence, the probability of producing offspring with the same genotype as in organism A is higher among relatives of this organism than among non-relatives. Thus, relatives have a higher value for the organism than non-relatives. Investment into relatives may be even more beneficial for self-production than investment into organism’s own progeny. In this case altruistic behavior may develop (Dawkins 1976).
Environment may have a considerable effect on the value of organisms because it changes the rate of survival and reproduction. Organism actions towards external objects become incorporated into the action-structure loop, and in this way they become parts of organization. As a result, an organism develops its Umwelt which associates external objects with specific actions.
Biological evolution leads to a gradual increase in complexity of Umwelts. Primitive organisms are unconscious investors because they don’t make any decisions. Their behavior is predetermined by their structure and environment. Investment strategy is improved (i.e., the value is increased) by differential survival and reproduction rather than by personal experience. Thus, their Umwelts are lineage-specific rather than organism-specific. Higher animals have individual perception of their values because they are able to distinguish between good and bad situations. Animals that evaluate situations can use previous experience to avoid bad situation in the future. As a result, they develop individual Umwelts which can be viewed as extensions of the lineage-Umwelt. External objects become linked to perceived values rather than to specific actions which makes individual Umwelts more flexible. For example, resources are associated with positive values whereas dangerous objects (e.g., enemies) have negative values. As a result, the animal evaluates the situation by examining its Umwelt and selects actions that can increase the value. The ability to evaluate situations changes the value of an organism because it helps to survive and reproduce. This creates a self-reference loop: an organism precepts its value but the value depends on perception.
Communication among living organisms develops only if it has value (Sharov 1992). Production of messages is beneficial only if these messages bring additional value to the organism, i.e., increase its self-production. For example, moth females emit pheromones and increase their probability of mating. This is the same kind of investment as production of eggs. Moreover, producing offspring can be considered as a sort of communication because an organism is a message (see above). The receiver never interprets the message unless it expects to increase its value after interpretation. Here I mean expectation in a broad sense including evolutionary (unconscious) expectation. Only in higher animals and humans expectation becomes conscious.
The ability to lie is often considered as an important feature of semiotic relationship (Sebeok 1976). Lie can be defined as a message that has a negative value for the receiver, but the receiver expects it to have a positive value. For example, some fireflies can imitate mating light signals of other firefly species on which they prey. The signal is an example of unconscious lie because it is fatal to the receiver who expects that it is beneficial.
The major trend in biological evolution is the increase of horizontal communication among simultaneously living organisms (Hoffmeyer 1997). This communication may result in a much faster self-production than vertical communication which is genetic heredity. Human evolution is mostly based on horizontal communication; thus reproduction of human bodies becomes less important than reproduction of habits and tools associated with these habits (Dawkins 1976, 1982). As a result, human values are associated with reproducing of life-styles rather than bodies.
The notion of value is essential for understanding semiosis because it specifies the relationship between the sign, interpreter, and interpretation. Interpretation is always an action that is expected to increase the value of the interpreter. It does not mean that interpreter’s value always increases after each interpretation. But in average, the value should increase because otherwise interpretation will not become a habit.
Metasystem Transition. The term ‘metasystem transition’ was suggested by Turchin (1977) for a process of developing new hierarchical levels. Turchin described a metasystem transition as follows: multiple copies of the original system are created followed by establishment of a new control mechanism that sets constraints on the behavior of all these copies. For example, multicellular organisms originated from individual cells that did not separate after reproduction. A new control mechanism was established at the multicellular level which regulated the size of a colony. Additional control was necessary for subsequent specialization of cells within a colony.
Metasystem transitions do not necessary occur at the highest hierarchical level. They may also happen at intermediate levels resulting in insertion of additional levels of control. For example, the complexity of the nervous system increased in evolution by duplication of neurons and their connections with subsequent establishment of new control levels over the set of neurons (Turchin 1977).
The theory of metasystem transition describes the process of organization growth in living systems but it does not capture evolutionary mechanisms that lead to duplication, aggregation and establishment of new controls. To some extent these mechanisms are considered in sociobiology which is targeted at explaining development of social behavior in animals (Wilson 1975). If social behavior is coded genetically and organisms live together with their close relatives, then this behavior may be favored by kin selection. However, sociobiology considers just one special mechanism of metasystem transition which is dangerous to extrapolate to other cases. For example, the cultural evolution of human society is definitely not based on changes in gene frequencies.
I think that the theory of metasystem transition should be integrated with the conception of value described above. Values exist at various hierarchical levels, and the major problem is to understand the correspondence between values at different levels. For example, cells may aggregate into a colony only if their individual values are increased after aggregation. The whole cannot exist without parts; thus, parts have some value for the whole. The evolutionary mechanism of metasystem transition should be very similar if not identical to economical mechanisms that lead to aggregation of people. Parts benefit from being in the whole and thus they can ‘pay taxes’ to the whole if these taxes are smaller than their benefits from being in the whole. This value excess (e.g., collected taxes) is used to perform the functions of the whole. According to the Hoffmeyer’s conception of a swarming body, the whole does not have its own ‘substance’. Thus, the whole can perform its functions only by delegating them to the parts. Parts receive additional value (e.g., salary) for performing the functions of the whole.
Turchin (1977) discussed metasystem transitions using the methodology of cybernetics and thus described the function of a higher-level system as ‘control’ of subsystems. Control means superiority of a system over its subsystems. It may involve feedback from subsystems but this feedback is defined by the higher-level system. If cybernetic methodology were applied to a human society, then it would be a totalitarian society in which the goal of the government is to keep control over its citizens using all available information.
Biosemiotics brings a new understanding of hierarchies: it does not imply the superiority of a system over its subsystems. Some systems which I call ‘strong’ are based on control (e.g., military organizations); but also there are ‘weak’ systems which mostly depend on their subsystems. For example, a club is a weak system because it depends on its members more than members depend on the club. People join the club voluntarily because by being in the club they increase their individual values. New hierarchical levels probably appear first as weak systems but then they may evolve towards stronger control. However, the evolution may go in the opposite direction as well. For example, human society was definitely evolving from strong systems (monarchies) to weak systems (democracies).
References
Calow, P. (1978). Life cycles: an evolutionary approach to the physiology of reproduction, development, and ageing. London: Chapman and Hall.
Conrad, M. (1983). Adaptability. The significance of variability from molecule to ecosystem. New York: Plenum Press.
Dawkins. R. (1976). The selfish gene. Oxford: Oxford University Press.
Dawkins, R. (1982). The extended phenotype: the long reach of the gene. San Francisco: Freeman.
Deely, J. N. (1982). Introducing semiotic: its history and doctrine. Bloomington: Indiana University Press.
Deely, J. N. (1992). Semiotics and biosemiotics: are sign-science and life-science coextensive? In Biosemiotics: The Semiotic Web 1991, T. A. Sebeok and J. Umiker-Sebeok (eds.), 47-75. Berlin: Mouton de Gruyter.
Dubois, D. and Prade, H. (1988). Possibility theory: an approach to computerized processing of uncertainty. New York: Plenum Press.
Eigen, M. and Winkler, R. (1981). Laws of the game: how the principles of nature govern chance. New York: Knopf: distributed by Random House.
Hoffmeyer, J. (1997). Signs of meaning in the Universe. Bloomington: Indiana University Press.
Gazzaniga, M. S. (1985). The social brain. New York: Basic Books.
Kampis, G. (1991). Self-modifying systems in biology and cognitive science: a new framework for dynamics, information, and complexity. New York: Pergamon Press.
Krampen, M. (1981). Phytosemiotics. Semiotica 36(3/4), 187-209.
Lotman, Y. M. (1990). Universe of the mind. A semiotic theory of culture. London: I. B. Taurus.
Margulis, L. and Sagan, D. (1991). Mystery dance: on the evolution of human sexuality. New York: Summit Books.
Minsky, M. L. (1986). The society of mind. New York: Simon and Schuster.
Pattee, H. H. (1972). The nature of hiererchical controls in living matter. pp. 1-22. In: Foundations of mathematical biology. Volume 1. Subcellular systems, R. Rosen, ed. New York, Acad. Press.
Pattee, H. H. (1995). Evolving self-reference: matter, symbols, and semantic closure. Communication and Cognition - Artificial Intelligence, 12(1-2), 9-28.
Pielou, E. C. (1974). Population and community ecology: principles and methods. New York: Gordon and Breach.
Rosen, R. (1967). Optimality principles in biology. New York: Plenum Press.
Sebeok, T. A. (1972). Perspectives in zoosemiotics. The Hague: Mouton.
Sebeok, T. A. (1976). Contributions to the doctrine of signs. Bloomington: Indiana University Press.
Sharov, A. A. (1992). Biosemiotics: a functional-evolutionary approach to the analysis of the sense of information. In Biosemiotics: The Semiotic Web 1991. Sebeok, T. A. and J. Umiker-Sebeok (eds.), 345-373. Mouton de Gruyter, Berlin.
Stepanov, Y. S. (1971). Semiotics. Moscow: Nauka (in Russian).
Turchin, V. F. (1977). The phenomenon of science. New York: Columbia University Press.
Uexkull, J. von 1940 (1982). The Theory of Meaning. Semiotica 42(1), 25-82.
Wiener, N. (1962). Cybernetics; or, control and communication in the animal and the machine. Cambridge (MA): M.I.T. Press.
Wilson, E. O. (1975). Sociobiology. Cambridge (MA): Belknap Press.
Zadeh, L. A. (1996). Fuzzy sets, fuzzy logic, and fuzzy systems: selected papers. River Edge, NJ: World Scientific.
Alexei Sharov 10/06/97