Jakob von Uexküll (1864-1944). Used by courtesy of the
J. v. Uexküll Center
|
A modified version of the paper is published: Sharov, A.A. 2001. Umwelt theory and pragmatism. Semiotica, Vol. 2001, Issue 134: 211-228." Other papers by Alexei Sharov Pragmatism and Umwelt-theoryAlexei SharovDepartment of Entomology, Virginia Tech, Blacksburg, Virginia 24061, sharov@vt.edu
1. Introduction |
Jakob von Uexküll (1864-1944). Used by courtesy of the
Jakob von Uexküll (1982) is known for his studies on how living organisms subjectively perceive and interpret their environments. The key notion of his theory is 'Umwelt' which is usually translated as subjective universe, although in German it simply means 'environment'. Most ecologists assume that all organisms in the ecosystem share the same environment, but Uexküll thought that organisms may have different Umwelts even if they live in the same place. A stem of a blooming flower is perceived differently by an ant, cicada-larva, cow, and human. Umwelt is not an ecological niche because niches are assumed to be objective units of an ecosystem which can be quantified using external measuring devices. On the contrary, Umwelt is subjective and is not accessible for direct measurement for the same reason that we have no direct access to perceptions of other people.
| |
|
The main idea of Uexküll is that each component of Umwelt has a functional meaning for an organism; it may be food, shelter, enemy, or simply an object that is used for orientation. An organism actively creates its Umwelt through repeated interaction with the world. It simultaneously observes the world and changes it; the phenomenon which Uexküll called a functional circle. The Umwelt-theory also implies that it is not possible to separate mind from the world (matter) because mind makes the world meaningful. The theory of meaning developed by Uexküll is coherent with the ideas of semiotics (theory of signs) proposed earlier by Charles Peirce (1955). Translating the theory of Uexküll into the language of semiotics, we can say that Umwelt is not a set of objects in the environment but rather a system of signs interpreted by an organism. Umwelt can be viewed as a language which is used mostly for internal communication within an organism or lineage. But in many cases it can be partially communicated with other organisms (lineages) via chemical, visual, and acoustic signals. But humans share a large portion of their Umwelts because of a highly developed horizontal communication system. The Umwelt-theory of Uexküll contradicts to the traditional positivistic methodology of science. The goal of science is viewed as the discovery of various aspects of the objective real world that exists unconditionally and independently from any observer, whereas the goal of the Umwelt-theory is to study the subjective universe of an organism. This contradiction brings us to the philosophical debate between realism and pragmatism. Pragmatists do not separate objects from interpreters, which is consistent with the Umwelt-theory. The debate between realism and pragmatism is important for developing the methodology of science and human knowledge in general. Below I describe several problems that are intrinsically related to this debate.
In this paper I suggest that Uexküll's Umwelt-theory provides a biological turn in pragmatism and semiotics. I will review major arguments pro and against biologically-grounded pragmatism, discuss the relationship between pragmatism and evolutionary theory, and finally show that the notion of reproductive value can be considered a universal measure of usefulness in intentional systems which may lead to the development of quantitative pragmatism in the future. 2. Pragmatism and SemioticsPragmatism is a philosophical theory developed by American thinkers Peirce (1955), James (1954), and Dewey (1998).  It is a monistic philosophy that assumes that all distinctions or boundaries are subjective (i.e., operational, instrumental, or conventional) including the boundary between the mind and matter. Subjectivity does not mean pure arbitrariness or randomness, instead it always has a component of self-interest or usefulness (hence the term 'pragmatism'). Because of usefulness, subjectivity is partially predictable. Usefulness implies activity that supports (or creates) existence (Dewey 1998). It also implies that structures and boundaries have a meaning for the system, because they perform functions that support the existence of the system. Pragmatism is organically related to semiotics, the theory of signs, founded by Peirce (1955). A sign is a triadic relationship between a sign vehicle that points to an object by invoking the interpretant (model of an object) in the head of the interpreter. 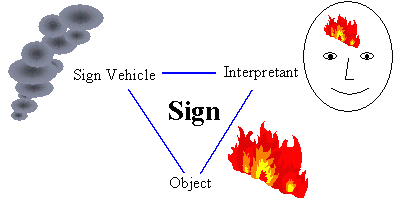 For example, smoke is a sign vehicle, fire is an object, and the idea of fire is an interpretant. Sign relationships are independent from physical relationships. They can be based on resemblance (icons), or conventions (symbols). Some signs (indexes) point to other objects via a physical relationship (e.g., position in space), but the nature of this relationship is not important for interpretation; it can be easily substituted by any other physical relationship. Semiotics and pragmatism are related in the following way. Both recognize an active and subjective role of an interpreter in its interaction with the world. Because all distinctions and boundaries are subjective, the world consists of signs rather than objects that exist independently from observers. Hence, semiotics describes the structure of the world. For solving some practical problems it may be convenient to consider matter and mind separately. But we need to keep in mind that this distinction is artificial; mind and matter are derivatives of something anterior to both, which can be called pure experience or firstness (Peirce 1955). Semiotics offered a novel understanding of human language. Linguistics was based on the Saussurean structuralistic view, according to which the meaning is determined by the structure of signs, both synchronic and diachronic. Peirce viewed structure as a product of evolution of signs. The meaning can be expressed in a particular structure, but the structure can be replaced by another structure without a change in the meaning. Pragmatism suggests to go beyond structure in a search for the meaning. Later, Morris (1971) suggested to distinguish 3 areas of semiotics: syntax, semantics, and pragmatics. Using this terminology, we can say that syntax and semantics were studied by Saussure, but it was Peirce who discovered pragmatics. 3. Uexküll's Biological Turn in PragmatismWhen Jacob von Uexküll developed his theory of meaning he knew neither Saussure not Peirce; thus, he developed his own terminology. But the general direction of his thought was very close to that of Peirce. The pragmatic nature of Uexküll's theory is seen from the following citation:
He considered usefulness the essential part of meaning. However, there are some important differences between Peirce and Uexküll. Being a biologist, Uexküll applied his version of semiotics to all living organisms including even individual cells. And he emphasized that the world perceived by various animals is entirely different. Peirce definitely believed that animals are capable of using signs, but most of his semiotics was oriented to humans. Thus, he did not realize that each species of animals live in a different subjective universe (Umwelt). Uexküll viewed usefulness as a biological adaptation which helps an organism to survive. Each sign is a part of one or several function circles that starts with a perception of a sign and ends with organism's action. He wrote:
Thus, signs are eventually interpreted as actions, whereas for Peirce, signs are interpreted mostly as mental concepts (interpretants). Animals classify things into categories that are associated with specific activities, e.g., as being 'drinkable', 'edible', 'sittable', etc. This is definitely a lower-level semiosis if compared with human language, but the nature of semiosis is easier to understand at this level. The most important feature of Uexküll's theory of meaning was the discovery that all signs used by an animal are organized into an integrated system, Umwelt, which represents the biological nature of this animal. Peirce did not have a comparable notion in his semiotics. He even did not include an interpreter into his triadic scheme of a sign, which indicates that an interpreter is de-emphasized. But it is the idea of 'self' that integrates signs into a subjective universe (Umwelt). In this question Uexküll definitely is ahead of Peirce. Umwelt-theory gives a new interpretation of space. Instead of a Newtonian absolute and transcendental space, Uexküll considers a functional space of an organism. Most biologists think that space exists independently from organisms that inhabit it. Uexküll viewed it differently: animals construct their own space by establishing relationships between meaning-carriers (i.e., signs). This does not mean that organisms do not interact; but an interaction is interpreted differently by each participant depending on its own model of space. The most primitive space is mapped on organism's body. Signals are identified according to the location of receptors on the body. More smart animals can identify objects at a distance; their space can be viewed as a set of objects that they know. Fast-moving animals can measure distance; hence they perceive space as a volume that contains various things. Perception of space is intrinsically related to the hierarchical structure of living organisms. Signs perceived by a sensor cell are transferred and then interpreted by the whole organism in a double way: as a location of that sensor cell in the organism (local sign) and as a content of the signal (content sign). Local signs are members of a class of signs which Uexküll (1982) called 'organizing signs'. Besides local signs, this class includes other hierarchical relationships: directional signs point the direction from one locality to another and require at least two participating cells; impulse-to-operation signs are signals from contracting muscles that are used by an organism to distinguish its own movements from movements of other things; and time signs are intrinsic clocks. Organizing signs represent the environment by coding it within organism's structure (location of sensors in the body). Thus, it is impossible to separate the structure of an organism from its Umwelt: 'However, the sky is a product of the eye� Eyeless living beings know neither sky nor sun.' (Uexküll 1982, p. 65). Organizing signs are opposed to signs of content that correspond to colors, sounds, smells, etc. Uexküll thought that the idea of 'objective world' is the result of abstraction from signs of content. Different Umwelts have a different content, but they remain identical in their structure. Uexküll viewed the body of a living organism as its tool. He wrote: 'Both animals and plants build living houses for themselves, i.e., their bodies, with whose help they lead their existence' (Uexküll 1982, p. 33). But if the body is a tool, then who is the user of this tool, where is the animal's self? The answer comes from a collective concept of self (Hoffmeyer 1996) which I will discuss later. Uexküll sees the hierarchy of living nature in both directions: down to individual cells, and up from organisms to species and finally to nature as a whole. He wrote: 'The basis of all life is the submergence of the ephemeral individuals for the sake of the long-lived species as a whole' (p. 60). For him, adaptation and usefulness are synonymous to harmony. His major metaphor for nature is an orchestra, in which individual organisms play their tunes. He wrote that processes in a growing organism follow a 'growth-melody'. Uexküll (1982) attempts to understand nature using musical laws of counterpoint. He wrote: 'Nature's techniques share common features with the creation of a work of art.' (p. 75). He viewed nature as a self-creating agent which assigns meaning to all its components. Uexküll's theory of nature and adaptation is close to vitalism (Driesch 1979) and nomogenesis (Berg 1969). The merit of these vitalistic theories is in a deep understanding of the unity of organisms. All of them viewed adaptations as specific relations between the structure of the body and organism's functions and behaviors. But the question about the origin of adaptations was not seriously discussed. At best these theories referred to a creative nature of life as a source of adaptations. The major competitor of vitalism was the Darwinian theory of natural selection that attempted to explain the origin of adaptations. All vitalists, including Uexküll, denied natural selection as a theory of adaptation. Uexküll (1982) thought that 'the removal of surplus individuals occurs in the interest of the balance of nature' (p. 61). Natural selection is interpreted by Darwinians as a passive sieve, i.e., as a mechanistic generator of adaptations that does not require any creative force. Darwinism views adaptation as something accidental ('blind variation') that was later picked by natural selection. Obviously, this interpretation is incompatible with vitalistic ideas of the unity of an organism. Vitalism had more in common with Lamarckian theory of evolution rather than with Darwinism. In summary, Uexküll grounded pragmatism and semiotics in biology. Each living system creates its own subjective universe (Umwelt) which consists of signs rather than objects. A sign is a product of organism's activity; it is represented by a functional circle which is useful for sustaining the existence of the organism. The idea that a sign is an adaptation integrates semiotics with biology a yields a new synthesis which is now known as biosemiotics (Sharov 1992, Hoffmeyer 1996). 4. The Meaning of ExistenceIn this section I compare pragmatism with logical positivism which remains the dominating philosophy of science. Logical positivism can be characterized by the following statements.
Positivism has a realistic approach towards facts (facts are 'stubborn'), and a phenomenological/pragmatic attitude towards theories (they are instruments to generalize facts). Facts are statements about existence of things which can be used to test various theories; but the existence itself in not disputed and does not require any explanation or justification. 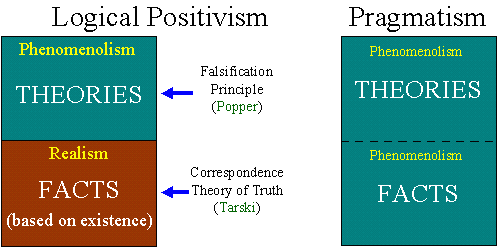 Pragmatism denies realism not only in the area of theories but also in the area of facts. Thus, there is no qualitative difference between facts and theories. Words that we use to formulate facts always have a hidden ambiguity, even in science (although in science, words are less ambiguous than in other areas of human discourse). Thus, it is impossible to describe any experiment exhaustively. |

For example, if we observe that water in a glass becomes frozen at 0� C, then we write in the journal: 'water becomes frozen at 0� C'. But this description does not capture all conditions of the experiment. May be there were some ions of Na+ in the water, or may be some hydrogen or oxygen atoms were isotopes. There are many other factors (e.g., gravity, vibration, radiation) that might affect the experiment. It is impossible to describe all these conditions with absolute accuracy. At best we can determine how inaccurate is our description. For example, we can say that concentration of other ions in the water was <0.001%.
| |
|
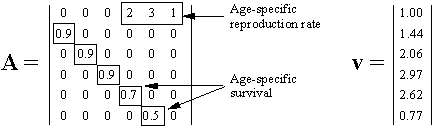
Because words are ambiguous, facts can not be communicated precisely; instead we describe them in a more or less generalized form, i.e., as theories. But may be facts can be communicated via objects themselves? For example, we can keep a sample of water and say that this water becomes frozen at 0� C. But how can we guarantee that this water remains the same? There is also ambiguity in the description of freezing. There may be thousands of various kinds of freezing that can not be expressed in words. Thus, facts can not be communicated precisely; they can be communicated only after some generalization which turns them into theories. The correspondence theory of truth fails because facts can not be communicated; only theories can be communicated. But theories can not be used to disprove other theories because they are questionable themselves. In contrast to positivism, pragmatism does not view existence as objective (i.e., observer-independent) reality. Instead, existence is evaluated subjectively according to its expected effects on the existence of other things in the future. For example, food exists for an animal because it increases chances of survival and reproduction (i.e., existence) of this animal. Subjective universe (Umwelt) of an animal consists of things that are useful in their life, e.g., food to consume, shelters to hide, and enemies to fear. Usefulness is used as a criterion of knowledge instead of truth. People wondered why the world around them is so useful and friendly. The answer is because humans have adapted to this world. A portion of animal's Umwelt can be communicated to neighbors via various kinds of signs (poses, smells, sounds, etc.). Another portion of Umwelt is communicated to offspring via genes. Humans share a large portion of their Umwelt because of intensive horizontal communication. But reality does not become objective even if it is shared. If I see a flying saucer in the sky, it becomes a part of my individual subjective reality. If I share this information with my neighbors and they also see the saucer, then it becomes a part of our shared subjective reality. There may be subjectivity at any level of the hierarchy: cells, individuals, families, nations, and even humanity as a whole. If the majority of scientific community accepts the existence of some phenomenon, this phenomenon does not become objective. But pragmatism does not imply that knowledge is impossible. On the contrary, it supports the growth of knowledge and especially scientific knowledge. But it supports knowledge not because it is true but because it is useful. Old geographic maps were not true but nevertheless they were useful. For a positivist, existence has no meaning; things simply exist as they are, period. Pragmatism sees the meaning of existence in its consequences. For example, James (1954) evaluated theories and beliefs according to their potential effect on human life and society. Human existence is also evaluated by its consequences in the future. If my life has no consequences, then I don't exist. The ethical principle of pragmatism is to create existence by caring about consequences and to bring meaning to life by doing this. In contrast, positivism is satisfied with a meaningless existence. The notion of 'existence for' is central for pragmatism because existence has meaning for some observer. It is opposed to a universal objective existence in positivism. The notion 'existence for' assumes the presence of a user, thus we need to answer the question who is the user. It appears that the user can not be separated from resources and tools that it uses. Our organs are all tools and resources; a hand is a tool, and fat reserves are resources (often unwanted). Then, where is the user, may be in the brain? But the brain is a tool too, it is personal computer for our body; and there is no homunculus inside our brain. Some people believe that there is immaterial soul that makes use of all body parts. This logic leads to objective idealism. But pragmatism does not need this assumption; a user is viewed as a collection of useful parts connected by useful relations.  Hoffmeyer (1996) used a metaphor of 'swarming body' to characterize the collective nature of an organism. Human consciousness also has a collective nature which was characterized by metaphors like 'the society of mind' (Minsky 1986) and 'swarm intelligence' (Hoffmeyer 1996). Applying the idea of 'existence for', we can say that an organism exists for itself because it is self-adapted. The body of an organism is a part of its Umwelt. Thus, 'existence for' may have two forms: (1) existence for something else, and (2) existence for itself. A system that exists for itself is an organism and things that exist for an organism are parts of its Umwelt. The major argument of positivism against pragmatism is that existing things are not necessary useful for observers. We use only a very small portion of all things that we observe. This argument points to one important aspect of usefulness that we have not discussed yet. Usefulness is hierarchical (or multiresolutional) both in time and space. Everybody knows the difference between short- and long-term usefulness. For example, by saving money in a retirement account you sacrifice your short-term needs for long-term benefits. Also, we distinguish values at the level of an individual, family, town, nation, and humanity as a whole. Human Umwelt consists of things that have values at various scales. It includes things that are utilized for our immediate needs (e.g., food), but it also includes things that have no immediate use, but may appear useful in the future. For example, most of our knowledge has no immediate use, but it increases our ability to solve various problems in the future. Useless and meaningless existence is an abstraction in which usefulness is ignored. But this abstraction is useful itself at a different level of hierarchy. Truth is a convenient substitution for usefulness but it works only until our model of the world remains useful. 5. Pragmatism and Evolutionary TheoryUexküll improved pragmatism by interpreting usefulness as a biological adaptation. But his criteria of adaptation (musical harmony, counterpoint) were anthropomorphic and qualitative. This is typical to vitalism and Lamarckism which simply refer to human intuition on what is useful and what is not. Quantitative criteria of usefulness are necessary to understand the emergence and evolution of adaptations. Darwinism explains the emergence of adaptations by the combination of inherited blind variation and natural selection. 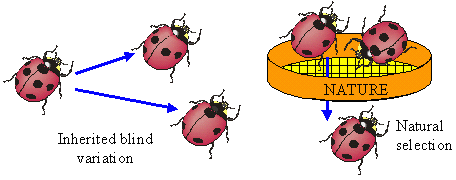 The advantage of Darwinism is that it offers a quantitative criterion of usefulness which is selective survival and reproduction. Although it looks attractive to combine pragmatism with the theory of natural selection, but the problem is that Darwinism is based on a mechanistic understanding of evolution which seems incompatible with pragmatism. Some biologists search for alternative evolutionary processes that may explain the origin of adaptations. For example, Kauffman (1993) argues that evolution is driven by two complimentary forces: natural selection and self-organization. By self-organization he means spontaneous structuring of systems via non-linear physical and chemical processes. He describes self-organization as 'order for free' implying that it does not require selection. But order does not mean adaptation; thus, Kauffman's theory does not offer an alternative explanation of adaptations. The theory of neutral evolution (Lewontin 1974) also does not explain the origin of adaptations. Studies of non-adaptive evolution may be important for some other purposes (e.g., for reconstruction of the phylogeny from DNA sequences) but they do not give quantitative criteria of usefulness. I believe that the contradiction between Darwinism and pragmatism results from erroneous interpretation of natural selection and there is no need to search for alternative evolutionary forces. Darwinism is not just a biological theory (which is not argued), it is also heavily loaded with positivistic philosophy. Positivism accepts only one true reality. If there are two different descriptions of a system and both are true, then either one of them can be derived from the other, or there is a third description from which both original descriptions can be derived. Thus, more detailed descriptions are more universal, and knowledge grows by adding details. Obviously, this strategy leads to reductionism. An organism is reduced to cells, and cells to molecules. But at the bottom level of reduction, there are only mechanical interactions of molecules. Thus, organisms are viewed as complex mechanisms, and creativity, intentionality, and free will are mere delusions. In contrast, pragmatism accepts multiple realities. Depending on how we interact with a system it may appear mechanistic or intentional. A system is intentional if it is more productive to interact with it as with intentional system rather than to consider it a mechanical system. A mechanism and organism are two metaphors that we use to understand reality.  Our experience indicates that the dynamics of a clock is better understood as a mechanism, and the behavior of a cat is better understood as an organism. However, this does not mean that for some purposes a cat can not be viewed as a biochemical machine. And the clock is not just a dumb machine. It can be viewed as a meme that reproduces and evolves. It may happen that some systems (e.g., the solar system, atom) appear to us as mechanisms simply because we have not found an appropriate spatial and temporal scales to see their intentionality. Thus, there is no such thing as being a mechanism. We view things as mechanisms if have not discovered their intentionality (but this does not mean that it is always productive to search for intentionality). Now we come to the central question: is it possible to find intentionality in the process of natural selection? The solution was suggested by Hoffmeyer (1996) who wrote that selection can be viewed as an intentional process at the level of lineages. Indeed, a lineage probes the environment by producing a variety of organisms and generating copies of those organisms that survived. Hoffmeyer (1996) suggested not to use the term 'natural selection' because the word 'natural' is often erroneously associated with nature as a super-agent that evaluates the fitness of organisms. Selection appears analogous to perception. Selective survival and reproduction of organisms plays the same role for a lineage as selective excitation of receptors for an organism (Sharov 1998). Animals can improve their behavior using vision which results from selective excitation of photoreceptors. If all photoreceptors become excited or none of them, then the animal can not use its vision to find food or to escape enemies. In the same way, a lineage can not evolve without sensing its Umwelt with selective survival and reproduction. 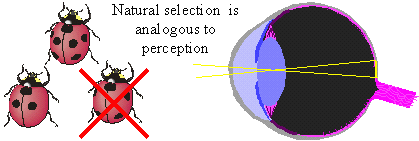 Perception requires information transfer. A photoreceptor sends an electric impulse to the brain. It is important that the brain recognizes impulses from individual photoreceptors. If nerve fibers become entangled, then the brain can not associate an incoming impulse with a specific receptor, and the information is lost. Heredity is the information transfer at level of lineages by which survived organisms transfer their genetic information to the next generation. If heredity were not enough accurate, then the information would be lost as in the case of entangled nerve fibers. There is a critical level of noise at which biological evolution becomes impossible; Eigen and Schuster (1979) called this phenomenon 'error catastrophe'. One of persistent confusions in Darwinism comes from the separation of heredity and variability, as if these two factors are entirely independent. Some portion of variability is indeed independent from heredity and can be 'intentionally' increased or decreased by a lineage. I will refer to this kind of variability as variability of freedom. However, there is another portion, which I will call the variability of error, which can not be separated from heredity. The variability of error corresponds to the resolution of perception (recognition limit). For example, the resolution of vision depends on the quality of optical characteristics of an eye and the size of photoreceptors. But when we draw a picture, we may intentionally blur the image and add some variability of freedom. The genetic variability of error is harmful, and living organisms reduce it by a variety of ways that include DNA proofreading, selective splicing, developmental constraints, apoptosis, immune responses, and others. Many diseases (e.g., cancer, mutations, chromosomal changes) result from error accumulation in transfer of biological information. On the contrary, the variability of freedom is useful (and even enjoyable for those organisms who have emotions). We are used to separate perception and action. First we see or hear, and then we act according to the information received (e.g., stop at the red light). But selective survival and reproduction of organisms is simultaneously a perception and action. We need to decide if this difference is important enough to drop the analogy between selection an perception. I believe that separation of perception and activity is artificial. Any perception involves some activity at least at a sub-cellular level. And activity is a response to some signal, i.e. it is a kind of perception. For example, muscle cells perceive incoming nerve impulses and contract in response. At the molecular level, the processes of perception and action are identical: they both include (1) an external impact that modifies molecular forces, and (2) subsequent change of the conformation of a molecule. Another possible argument against the analogy between perception and selection is that organisms are able to perceive negative information whereas lineages are not able. The absence of a signal can be a signal for an organism, in other words, our nervous system can use the negation operator. Lineages have no negation operator because they do not notice that some organisms are missing. This idea is embedded in the metaphor of 'blind variation' which is widely used by Darwinism. No matter how many organisms have died because of some particular mutation, the lineage will continue producing this mutation in the future; it does not learn from errors. I have two arguments in defense of the analogy between perception and selection. First, the negation operator is a result of reconstruction (generalization) rather than perception, and second, lineages can use negation by generalizing sequential information. We see a black spot on a white background despite the fact that our receptors do not get any photons coming from this black spot. The photoreceptors, on which the black spot is projected in the eye, do not send any impulses to the brain. However, our brain reconstructs the black spot and presents it to our consciousness as an object. This reconstruction requires generalization of information collected from multiple receptors; it would be impossible to see a black spot if we had just one photoreceptor. 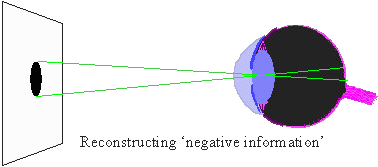 The absence of a signal from the receptor may have two possible reasons: (1) there is no stimulus, and (2) the receptor is not functional (e.g., it died). The brain assumes that the receptor it functional but this assumption may be wrong. The brain also knows that there are no photoreceptors in the 'blind spot' area. We don't see any black spot there although the brain does not get any signals from that area in the retina. Thus, negative information is not perceived by an organism but it is reconstructed using a particular model of vision. Let us see if anything comparable exists in evolution. The metaphor of 'blind variation' is the favorite one among Darwinists. Blind variation means that characteristics of an organism may change randomly in its progeny. However, this term is never applied to stable characteristics that are specific for higher taxons. Nobody expect foxes to produce randomly rabbits, cockroaches, and frogs (although, Lysenko in Russia expected that rye can produce wheat). These characteristics are stable because organisms have developmental constraints that prevent their change. The importance of these constraints was explained by the theory of stabilizing selection (Shmalgauzen 1949). Later, the same arguments were repeated by Conrad (1983) and Kauffman (1993). Constraints for variation are not mechanistic as assumed by neo-Darwinism, but semiotic because lineages have developed them in evolution to decrease the variation of errors. Organisms know the difference between good and bad changes, and this knowledge arose from their experience in previous generations. This means that lineages do not repeat their errors indefinitely as assumed by the 'blind variation' metaphor. Finally they learn how to avoid errors by creating proofreading mechanisms and developmental constraints, in other words, they reconstruct negative information. Let us consider a lineage in a narrow evolutionary nich (see figure below). If a lineage does not reconstruct negative information it will continue producing the same variation. However, if the proofreading is improved via stabilizing selection, then the variation will decrease with time. This means that the lineage 'realises' the bounds of its evolutionary niche. 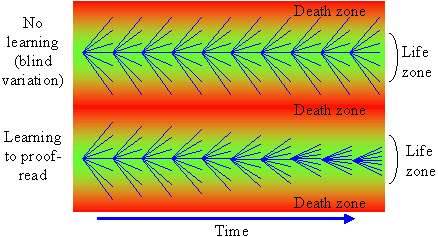 But the reconstruction of negative information is different in visual perception and selective survival and reproduction. Visual reconstruction is mostly parallel, i.e., it is based on generalization of visual signals simultaneously arriving from multiple receptors. On the contrary, lineages can generalize only sequential information (here I ignore sexual reproduction which adds parallel signals). The existence of a lineage means that it was successful in avoiding errors in past generations. Its genetical and developmental constraints represent the rules by which errors can be avoided. This does not mean that violation of these rules is always harmful, but the lineage assumes that it is harmful (similarly, some people believe that anything new is bad). Thus, lineages have something like consciousness, but it is extremely slow because they can generalize only sequential information. It takes millions of generations for them to learn how to avoid a single developmental error. Sexual reproduction can accelerate this process because an organism obtains access to the genetically-recorded experience of other organisms and not just of its predecessors. Because natural selection is equivalent to perception, lineages are intentional systems. In fact, the intentionality of individual organisms originated from the intentionality of lineages. A multicellular organism is a lineage of cells that live together and cooperate. As a result, natural selection of whole organisms evolves into selection of cells or even molecules at sub-organismal levels. With selection at sub-organismal level there is no need to waste the whole body in order to get 1 bit of information. Selection at sub-system levels take various forms: selective binding of molecules, selective interaction between neurons (Edelman 1987), and selective reproduction of lymphocytes in the immune system (Golub 1981). Thus, pragmatism can be integrated with the theory of natural selection if it is re-interpreted as perception at the level of lineages. Selection is the essential component of perception because information is always acquired via selective response of system components. Evolution is similar to learning, both are intentionally-driven processes that increase usefulness (fitness). 6. Umwelt, the World of ValuesValue is important for pragmatism because it is the quantitative measure of usefulness. Value is often considered a non-scientific concept because it is subjective and non-measurable. I would argue that values can be measured and studied scientifically despite their subjectivity. Economists measure the present value, PV, of investment as a sum of inflation-corrected net returns, V(t), expected in the future: 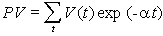 In a linear model of population growth, the reproductive value of organisms is equal to the left eigenvector of the matrix that describes population dynamics. For example, in the model of Leslie (19), the state of a population is characterized by the vector of age distribution, xt, that shows the number of organisms in each age group at time t. The population dynamics is described by equation xt +1 = A xt where A is the Leslie matrix (Fig. 1)., then the vector of reproductive values, v, is the left eigenvector of matrix A. Newborn organisms have a reproductive value =1 (Fig. 1). As they mature, their reproductive value increases to 2.97, and then decreases to 0.77 in the oldest age group. Vector v satisfies the equation ATv = lv, where AT is the transposed matrix A, and l is the largest real eigenvalue, which is the rate of population increase (l = 1.293 for the matrix in Fig. 1). Computation of reproductive values in non-linear systems is more complicated because these systems may have limit cycles or chaotic dynamics (9).
A Leslie matrix A and the vector of reproductive values v. If the dynamics of the system is non-linear, then the time scale of the model should be increased until the dynamics will become close to linear. The relative rate of value production in the population is equal to fitness (real positive eigenvalue). If a mutation increases the average relative rate of value production, then the frequency of this mutation will increase because of natural selection. Thus, an organism can be viewed as an investor whose profit is self-production (Sharov 1992). To maximize the rate of self-production, it should produce systems with maximal total value. For example, an adult insect can immediately spend its entire energy on laying eggs and then die, or it may lay a few eggs and continue living. If the decrease in the value of this insect after laying an egg is smaller than the value of an egg, then it is beneficial to continue laying eggs. However, if the decrease in the value of adult insect is greater than the value of an egg, then it is beneficial to stop laying eggs because it is better to postpone egg production. Selective survival and reproduction is the only conceivable way by which the adaptive evolution could start. However, on later stages of evolution, actual reproduction can be replaced partially by virtual reproduction by which I mean using models and simulations rather that real organisms for testing the performance of the system. Campbell (1988) called it 'vicarious selection' which replaces natural selection. For example, animals can estimate the value of their actions before natural selection takes place. Thus, they are able to optimize their behavior faster than organisms that rely solely on natural selection. Communication among living organisms develops only if it has value (Sharov 1992). Production of messages is beneficial only if these messages bring additional value to the organism, i.e., increase its self-production. For example, moth females emit pheromones and increase their probability of mating. This is the same kind of investment as production of eggs. Moreover, producing offspring can be considered as a sort of communication because an organism is a genetical message. The receiver never interprets the message unless it expects to increase its value after interpretation. Here I mean expectation in a broad sense including evolutionary expectation. Only in higher animals and humans expectation becomes conscious. Values exist relative to a particular quasi-species (Eigen and Schuster 1979) which consists of one or several components with stable relative frequencies. If a quasi-species changes in evolution, its reproductive values change too. For example, if an organism develops effective protection against natural enemies, then the relative value of an egg increases because there are greater chances to survive to the adult stage. As the value of an egg increases, it becomes beneficial for a parent to invest additional resources into a single egg. Humans can identify themselves with various quasi-species depending on their life goals. They may see the meaning of their existence in biological survival and reproduction, capital growth, dissemination of their ideas, etc. Their values are determined by the quasi-species selected. The value of a part or process in a system can be measured by the contribution of that part or process to the value of the whole system. The value of a system is always greater than the sum of values of its parts because interactions between parts generate additional value. The integrity of a system can be measured by the difference between its value and the sum of values of its parts. The body is the collection of most valuable parts and processes. Thus, the boundary of the system is determined by the value of its parts. A conflict is a situation when one part has value for two different systems. These conflicts are resolved either by aggression or cooperation which is exchange of values. In hierarchical systems, values exist at multiple structural levels and time scales. For example, in the human society, there are values at the level of individuals, families, nations, religions, and humanity as a whole. Usually, values of higher structural levels are achieved at larger time scales. At each level of hierarchy, sub-systems develop level-specific dynamical models which determine their values. System boundaries depend on values because an element belongs to the system if it has a value We need to develop a new system of terms to speak about values. For example, the term 'action' is often applied to non-intentional objects (e.g., the bomb exploded). I suggest to use this term only for intentional subjects. It is a human who exploded the bomb (if it was intentional). Otherwise, is was not an action but an accident. The same confusion happens if the term 'function' is applied to mechanisms. I suggest to reserve the term 'function' only for those processes that generate value for some quasi-species. For example, computation is not a function of the computer, it is rather a human function that requires a computer. Two important classes of functions are control and freedom. Control is a stabilizing function and freedom is a diversifying function. Homeostasis is a specific kind of control which creates an equilibrium point for system dynamics. In biological, social, and technical evolution, control and freedom develop side by side. Any increase in freedom may have harmful consequences; thus, control mechanisms become developed to restrict (regulate) the use of freedom. At the beginning of life, control mechanisms were weak and there was a danger to loose information due to error catastrophe (Eigen and Schuster 1979). To avoid error catastrophe, pre-biological systems should have very limited freedom so that freedom and control are in balance. Better control stimulates the growth of freedom, and growth of freedom stimulates better control. This positive feedback is the engine of biological and social progress. References
Alexei Sharov 11/23/99 |