Age-dependent life-tables
Age-dependent life table shows organisms' mortality (or survival) and reproduction rate (maternal frequency) as a function of age. In nature, mortality and reproduction rate may depend on numerous factors: temperature, population density, etc. When building a life-table, the effect of these factors is averaged. Only age is considered as a factor that determines mortality and reproduction.Example. Consider a sheep population which is censused once a year immediately after breeding season:
|
Age, years (x) | Probability of surviving to age x (lx) | No. of female offspring born to a mother of age x (mx) |
| 0 | 1.000 | 0.000 |
| 1 | 0.845 | 0.045 |
| 2 | 0.824 | 0.391 |
| 3 | 0.795 | 0.472 |
| 4 | 0.755 | 0.484 |
| 5 | 0.699 | 0.546 |
| 6 | 0.626 | 0.543 |
| 7 | 0.532 | 0.502 |
| 8 | 0.418 | 0.468 |
| 9 | 0.289 | 0.459 |
| 10 | 0.162 | 0.433 |
| 11 | 0.060 | 0.421 |
Survivorship curves
Survival probabilities lx are often plotted against age x. These graphs are called "survivorship curves". They show, at what age death rates are high and low. The following graphs show two survivorship curves for domestic sheep (data from Caughley 1967) and for lapwings or green plovers (Vanellus vanellus) in Britain (data from Deevey 1947):
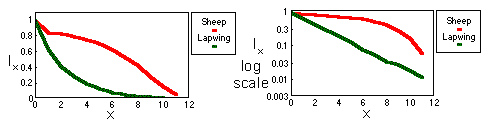
Survivorship curve is exponential (with negative growth) for the lapwing. This means that survival rate is independent of age. In the log-scale, survivorship curve becomes a straight line (see above).
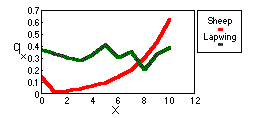
Age-specific mortality is estimated using equation:
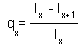
Sheep mortality generally increases with age; and the slope of survivorship curve becomes steeper to the end. Humans have a similar shape of survivorship curve.
Characteristics derived from life-tables
Net reproductive rate, R0, is the average number of female offspring born to a sheep considered at age 0. Consider N new-born sheep. Some of them will die without producing any offspring (zero offspring), others will produce one or several offspring. R0 is the average number of female offspring produced in the entire group of N sheep.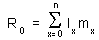
In our example, R0 = 1×0 + 0.845×0.045 + 0.824×0.391 + ... = 2.513. It means that an ewe produces 2.513 ewe lambs in average in a lifetime.
Average generation time, T, is estimated using equation:
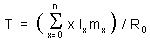
In our example, T = (0×1×0 + 1×0.845×0.045 + 2×0.824×0.391 + ...) / 2.513 = 12.825 / 2.513 = 5.1 years. It means that the average age of mothers when they give birth to an eve lamb is 5.1 years.
Note: If organisms breed continuously, then generation time will be overestimated using this equation because all births are summed over the period between census dates which is equal to one age step. Generation time can be adjusted by subtracting half of age step.
Now it is possible to estimate approximate value of intrinsic rate of increase r using the following logic. We assume discrete generations with generation time T=5.1 years and net reproductive rate of R0 =2.513. If population size at zero time was N0, then after T years, the population will grow to NT = N0×R0. According to an exponential model,
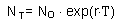
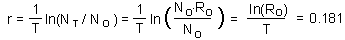
where ln is natural logarithm (logarithm with base e =2.718). We got the equation for r:
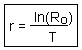
Note: This is an approximate estimation of r because we used a simplified assumption that generations are discrete. Accurate estimation of r will be discussed in the next chapter.
Time units used for age measurement
If survival and reproduction are continuous processes without any cyclic change, then any time units may be suitable: days, weeks, months, years. Time units should provide sufficient resolution. For example, if the life span of an organism is 2 months, then taking 1 month as a unit will result in only two age intervals which is definitely not sufficient. In most cases, the number of age intervals is in the range from 10 to 50.If survival or reproduction are cyclic (e.g., seasonal), then one cycle can be taken as a time unit. In this case it may happen that the number of age intervals will drop to 2 or 3. However, it may be not very dangerous if reproduction is limited to a short period within the year because there will be little age difference between organisms born in the same year.
If survival and reproduction are cyclic but the entire life span is less or equal to this cycle, then time units should be smaller than the cycle length. Age-dependent life-tables can be built for the entire population only if the breeding period is short and therefore organisms' development is synchronized. Otherwise, separate life-tables should be built for subpopulations that start their development at different seasons.
Determining survival of organisms till age x (lx)
For domestic animals or for populations reared in the laboratory, it is possible to observe the fate of a large group of individuals that all started life simultaneously. Survivors can be counted at regular time intervals and lx values can be easily estimated. Similar technique can be applied to non-moving organisms (plants, sedimentary animals). It is possible to mark a large number of individuals and to trace their fate.This kind of analysis is usually impossible in populations of moving organisms. There are two principal methods to deduce lx values in this case: by determining the age distribution in the population, or by determining the ages at death.
If the population is stationary (i.e., population numbers and age distribution do not change) then the number of new-born organisms x time units ago was the same as now; and survivors of that group of organisms are of age x. Thus,
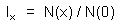
where N(x) is the number of organisms of age x. Here we assumed that age can be accurately measured. In many species the number of "growth rings" in specific organs is equal to age in years. Examples of such organs are: stems of trees, scales of fishes, horns in sheep, roots of canine teeth in bears, otholits in fishes. The weight of the eye lens can be used for age measurement in some animal species. However, in many populations, measuring age is a difficult problem.
Assumption of population stationarity usually is taken a-priori if no historical data exist. In cases when age distribution has a periodic component, we can speculate that it resulted from fluctuations in population numbers. Periodic component can be filtered out using regression methods.
Consider a large number of carcasses whose age have been determined. We assume that the probability of detecting a carcass does not depend on the age of animal at death. The proportion of individuals that were at age x when they died is dx. These individuals survived to age x but did not survive to age x+1. Thus,
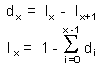
The method of estimation of lx from age of carcasses also requires stability of population numbers and of age structure.
Determining reproduction rates (mx)
Reproduction rate (=maternal frequency) is equal to the number of female offspring produced per one mother in age interval from x to x+1. Survival of mothers and offspring during this time interval should be included into m ; i.e., mx is equal to the total number of offspring produced in one time interval and survived till the end of this period divided by the initial number of parent females at the beginning of the time interval.Reproduction rates are easy to determine in laboratory reared animals or plants. In natural populations of mammals, maternal frequencies can be derived from the proportion of pregnant and/or lactating animals. In birds, maternal frequencies can be determined from the average number of chick per nest. Indirect measures of maternal frequencies should be used with caution because they may be biased.